

Мы поможем в написании ваших работ!
ЗНАЕТЕ ЛИ ВЫ?
|
Тема 14. 1. Метаболизм эритроцитов
Содержание книги
- Регуляция активности пируваткиназы в печени осуществляется путем фосфорилирования (дефосфорилирования) в зависимости от ритма питания
- Структурная организация межклеточного матрикса (суставной хрящ, базальные мембраны, субэпителиальные слои)
- Гликозилирование лизина под действием гликозилтрансфераз прекращается по мере формирования трехспиральной структуры.
- Патологий, связанных с уменьшением эластичности сосудов. При недостаточной активности металлопротеиназ развивается фиброз тканей и неадекватный иммунный ответ.
- Хондроитинсульфат; 2 - кератансульфат; 3 - коровый белок; гк - гиалуроновая кислота
- Тема 7. 5. Структурная организация межклеточного матрикса (суставной хрящ, базальные мембраны, субэпителиальные слои)
- Регуляция процесса. Амф, гмф, имф, ди- и трифосфаты адениловых и гуаниловых нуклеотидов ингибируют ключевые реакции своего синтезааллостерически по механизму отрицательной обратной связи.
- Частым нарушением катаболизма пуринов является гиперурикемия, которая возникает, когда в плазме крови Концентрация мочевой кислоты превышает норму.
- Синтез цтф из утф осуществляет цтф-синтетаза, используя амидную группу глн и энергию атф для аминирования пиримидинового кольца.
- Тема 10. 3. Биосинтез дезоксирибонуклеотидов.
- Тема 10. 4. Механизмы действия противовирусных и противоопухолевых препаратов на ферменты синтеза рибо- и дезоксирибонуклеотидов
- И рецепторов. Особенно много сфинголипидов в нервной ткани, где они формируют миелиновые оболочки нейронов.
- Тема 8. 3. Хиломикроны - транспортная форма экзогенных жиров
- Модульная единица 2 биосинтез высших жирных кислот и жиров
- Реакции восстановления обеспечивают синтез насыщенного алифатического радикала жирных кислот
- Запасание жиров в жировой ткани - так называемое депонирование жиров - происходит в абсорбтивный период, когда увеличивается соотношение инсулин - глюкагон.
- Первичное ожирение развивается в результате алиментарного дисбаланса - избыточной калорийности питания по сравнению с расходами энергии.
- Модульная единица 3 жиры, жирные кислоты и кетоновые тела как источники энергии. Эйкозаноиды, Строение, синтез и биологические функции
- Скелетные мышцы и почки используют кетоновые тела даже при их низкой концентрации в крови.
- Выведение кетоновых тел, в том числе и ацетона, с мочой (кетонурия), потом выдыхаемым воздухом является способом выведения избытка кетоновых тел из организма и уменьшения таким образом ацидоза.
- Ацетильный остаток переносится с молекулы аспирина на oh-группу фермента и необратимо ингибирует его
- Модульная единица 4 обмен холестерола, его регуляция и транспорт кровью. Дислипопротеинемии. Биосинтез и функции желчных кислот. Желчнокаменная болезнь
- В поддержании гомеостаза холестерола в организме. Биохимия желчнокаменной болезни
- Тема 8. 14. Роль липопротеинов в транспорте
- Атеросклероз представляет собой заболевание, при котором поражается внутренний слой артерий за счет отложения холестерола в интиме сосудов.
- Тема 9. 2. Переваривание белков в желудке и кишечнике, всасывание аминокислот
- Тема 9. 3. Трансаминирование и дезаминирование аминокислот
- Тема 9. 4. Обмен аммиака: источники, превращение в тканях
- Тема 9. 5. Орнитиновый цикл и его биологическая роль
- Тема 9. 6. Гипераммониемия и ее причины
- Тема 9. 8. Биосинтез заменимых аминокислот
- Тема 9. 10. Обмен метионина. Реакции трансметилирования
- Тема 9. 11. Обмен фенилаланина, тирозина и гистидина в разных тканях
- Тема 9. 12. Заболевания, связанные с нарушением обмена фенилаланина и тирозина
- Тема 9. 13. Биогенные амины: синтез, инактивация, биологическая роль
- Тема 11. 1. Роль гомонов в регуляции метаболизма
- Тема 11. 3. Строение и биосинтез гормонов
- Тема 11. 4. Регуляция обмена основных энергоносителей при нормальном ритме
- Тема 11. 5. Изменения метаболизма при гипо- и гиперсекреции гормонов
- Тема 11. 6. Изменения гормонального статуса и метаболизма при голодании и физической работе
- Тема 11. 7. Изменения гормонального статуса и метаболизма при сахарном диабете
- Тема 11. 8. Регуляция водно-солевого обмена
- Тема 11. 9. Регуляция обмена кальция и фосфатов. Строение, синтез и механизм действия паратгормона, кальцитриола и кальцитонина
- Модуль 12 обезвреживание токсических веществ в печени
- Тема 12. 2. Обезвреживание продуктов катаболизма аминокислот в кишечнике
- Тема 12. 3. Биотрансформация лекарств
- Тема 12. 5. Химический канцерогенез
- Тема 13. 1. Синтез гема и его регуляция
- Тема 14. 1. Метаболизм эритроцитов
- Тема 14. 2. Особенности метаболизма фагоцитирующих клеток
Похожие статьи вашей тематики
Эритроциты - высокоспециализированные клетки, которые переносят кислород от легких к тканям и диоксид углерода, образующийся при метаболизме из тканей к альвеолам легких. В результате дифференцировки эритроциты теряют ядро, рибосомы, митохондрии, эндоплазматический ретикулум. Эти клетки имеют только плазматическую мембрану и цитоплазму. Они не содержат ядра, поэтому неспособны к самовоспроизведению и репарации возникающих в них повреждений. Двояковогнутая форма эритроцитов имеет большую площадь поверхности по сравнению с клетками сферической формы такого же размера. Это облегчает газообмен между клеткой и внеклеточной средой. Вместе с тем такая форма и особенности строения
цитоскелета и плазматической мембраны обеспечивают большую пластичность эритроцитов при прохождении ими мелких капилляров.
Метаболизм глюкозы в эритроцитах представлен анаэробным гликолизом и пентозофосфатным путем превращения глюкозы. Эти процессы обусловливают сохранение структуры и функций гемоглобина, целостность клеточной мембраны и образование энергии для работы ионных насосов.
1. Гликолиз обеспечивает энергией работу транспортных АТФаз, а также протекающие с затратой АТФ гексокиназную и фосфофруктокиназную реакции гликолиза. NADH, образующийся в ходе анаэробного гликолиза, является коферментом метгемоглобинредуктазы, катализирующей восстановление метгемоглобина в гемоглобин. Кроме того, в эритроцитах присутствует фермент бисфосфоглицератмутаза, превращающий промежуточный метаболит этого процесса 1,3-бисфосфоглицерат в 2,3-бисфосфоглицерат. Образующийся только в эритроцитах 2,3-бисфосфоглицерат служит важным аллостерическим регулятором связывания кислорода с гемоглобином. На окислительном этапе пентозофосфатного пути превращения глюкозы образуется NADPH, участвующий в восстановлении глутатиона. Последний используется в антиоксидантной защите эритроцитов (рис. 14.1).
 Рис. 14.1. Образование и обезвреживание активных форм кислорода в эритроцитах: Рис. 14.1. Образование и обезвреживание активных форм кислорода в эритроцитах:
1 - источник супероксидного аниона в эритроцитах - спонтанное окисление Fe2+ в геме гемоглобина; 2 - супероксиддисмутаза превращает супероксидный анион в пероксид водорода и О2; 3 - пероксид водорода расщепляется каталазой или глутатионпероксидазой; 4 - глутатионредуктаза восстанавливает окисленный глутатион; 5 - на окислительном этапе пентозофосфатного пути превращения глюкозы образуется NADPH, необходимый для восстановления глутатиона; 6 - в глицеральдегидфосфатдегидрогеназной реакции гликолиза образуется NADH, участвующий в восстановлении железа метгемоглобина метгемоглобинредуктазной системой
2. Большое содержание кислорода в эритроцитах определяет высокую скорость образования супероксидного анион-радикала O2-, пероксида водорода Н2О2 и гидроксил-радикала ОН'.
Постоянным источником активных форм кислорода в эритроцитах является неферментативное окисление железа гемоглобина:
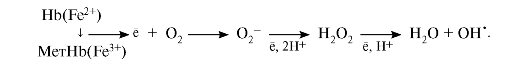 Активные формы кислорода могут вызвать гемолиз эритроцитов. Эритроциты содержат ферментативную систему, предотвращающую токсическое действие радикалов кислорода и разрушение мембран эритроцитов. Активные формы кислорода могут вызвать гемолиз эритроцитов. Эритроциты содержат ферментативную систему, предотвращающую токсическое действие радикалов кислорода и разрушение мембран эритроцитов.
3. Нарушение любого звена ферментативной системы обезвреживания активных форм кислорода приводит к снижению скорости этого процесса. При генетическом дефекте глюкозо-6-фосфатдегидрогеназы и приеме некоторых лекарств, являющихся сильными окислителями, потенциал глутатионовой защиты может оказаться недостаточным. Это приводит к повышению содержания в клетках активных форм кислорода, вызывающих окисление SH-групп молекул гемоглобина. Образование дисульфидных связей между протомерами гемоглобина и метгемоглобина приводит к их агрегации - образованию телец Хайнца (рис. 14.2).
 Рис. 14.2. Схема образования телец Хайнца - агрегации молекул гемоглобина. Рис. 14.2. Схема образования телец Хайнца - агрегации молекул гемоглобина.
В норме супероксиддисмутаза катализирует образование пероксида водорода, который под действием глутатионпероксидазы превращается в Н2О. При недостаточной активности ферментов обезвреживания активных форм кислорода происходит окисление SH-групп в остатках цистеина протомеров метгемоглобина и образование дисульфидных связей. Такие структуры называются тельцами Хайнца
Последние способствуют разрушению эритроцитов при попадании их в мелкие капилляры. Активные формы кислорода, вызывая перекисное окисление липидов мембран, разрушают мембраны.
|




