Мы поможем в написании ваших работ!
ЗНАЕТЕ ЛИ ВЫ?
|
Тема 9. 10. Обмен метионина. Реакции трансметилирования
Содержание книги
- Регуляция активности пируваткиназы в печени осуществляется путем фосфорилирования (дефосфорилирования) в зависимости от ритма питания
- Структурная организация межклеточного матрикса (суставной хрящ, базальные мембраны, субэпителиальные слои)
- Гликозилирование лизина под действием гликозилтрансфераз прекращается по мере формирования трехспиральной структуры.
- Патологий, связанных с уменьшением эластичности сосудов. При недостаточной активности металлопротеиназ развивается фиброз тканей и неадекватный иммунный ответ.
- Хондроитинсульфат; 2 - кератансульфат; 3 - коровый белок; гк - гиалуроновая кислота
- Тема 7. 5. Структурная организация межклеточного матрикса (суставной хрящ, базальные мембраны, субэпителиальные слои)
- Регуляция процесса. Амф, гмф, имф, ди- и трифосфаты адениловых и гуаниловых нуклеотидов ингибируют ключевые реакции своего синтезааллостерически по механизму отрицательной обратной связи.
- Частым нарушением катаболизма пуринов является гиперурикемия, которая возникает, когда в плазме крови Концентрация мочевой кислоты превышает норму.
- Синтез цтф из утф осуществляет цтф-синтетаза, используя амидную группу глн и энергию атф для аминирования пиримидинового кольца.
- Тема 10. 3. Биосинтез дезоксирибонуклеотидов.
- Тема 10. 4. Механизмы действия противовирусных и противоопухолевых препаратов на ферменты синтеза рибо- и дезоксирибонуклеотидов
- И рецепторов. Особенно много сфинголипидов в нервной ткани, где они формируют миелиновые оболочки нейронов.
- Тема 8. 3. Хиломикроны - транспортная форма экзогенных жиров
- Модульная единица 2 биосинтез высших жирных кислот и жиров
- Реакции восстановления обеспечивают синтез насыщенного алифатического радикала жирных кислот
- Запасание жиров в жировой ткани - так называемое депонирование жиров - происходит в абсорбтивный период, когда увеличивается соотношение инсулин - глюкагон.
- Первичное ожирение развивается в результате алиментарного дисбаланса - избыточной калорийности питания по сравнению с расходами энергии.
- Модульная единица 3 жиры, жирные кислоты и кетоновые тела как источники энергии. Эйкозаноиды, Строение, синтез и биологические функции
- Скелетные мышцы и почки используют кетоновые тела даже при их низкой концентрации в крови.
- Выведение кетоновых тел, в том числе и ацетона, с мочой (кетонурия), потом выдыхаемым воздухом является способом выведения избытка кетоновых тел из организма и уменьшения таким образом ацидоза.
- Ацетильный остаток переносится с молекулы аспирина на oh-группу фермента и необратимо ингибирует его
- Модульная единица 4 обмен холестерола, его регуляция и транспорт кровью. Дислипопротеинемии. Биосинтез и функции желчных кислот. Желчнокаменная болезнь
- В поддержании гомеостаза холестерола в организме. Биохимия желчнокаменной болезни
- Тема 8. 14. Роль липопротеинов в транспорте
- Атеросклероз представляет собой заболевание, при котором поражается внутренний слой артерий за счет отложения холестерола в интиме сосудов.
- Тема 9. 2. Переваривание белков в желудке и кишечнике, всасывание аминокислот
- Тема 9. 3. Трансаминирование и дезаминирование аминокислот
- Тема 9. 4. Обмен аммиака: источники, превращение в тканях
- Тема 9. 5. Орнитиновый цикл и его биологическая роль
- Тема 9. 6. Гипераммониемия и ее причины
- Тема 9. 8. Биосинтез заменимых аминокислот
- Тема 9. 10. Обмен метионина. Реакции трансметилирования
- Тема 9. 11. Обмен фенилаланина, тирозина и гистидина в разных тканях
- Тема 9. 12. Заболевания, связанные с нарушением обмена фенилаланина и тирозина
- Тема 9. 13. Биогенные амины: синтез, инактивация, биологическая роль
- Тема 11. 1. Роль гомонов в регуляции метаболизма
- Тема 11. 3. Строение и биосинтез гормонов
- Тема 11. 4. Регуляция обмена основных энергоносителей при нормальном ритме
- Тема 11. 5. Изменения метаболизма при гипо- и гиперсекреции гормонов
- Тема 11. 6. Изменения гормонального статуса и метаболизма при голодании и физической работе
- Тема 11. 7. Изменения гормонального статуса и метаболизма при сахарном диабете
- Тема 11. 8. Регуляция водно-солевого обмена
- Тема 11. 9. Регуляция обмена кальция и фосфатов. Строение, синтез и механизм действия паратгормона, кальцитриола и кальцитонина
- Модуль 12 обезвреживание токсических веществ в печени
- Тема 12. 2. Обезвреживание продуктов катаболизма аминокислот в кишечнике
- Тема 12. 3. Биотрансформация лекарств
- Тема 12. 5. Химический канцерогенез
- Тема 13. 1. Синтез гема и его регуляция
- Тема 14. 1. Метаболизм эритроцитов
- Тема 14. 2. Особенности метаболизма фагоцитирующих клеток
Похожие статьи вашей тематики
1. Метионин - незаменимая аминокислота, необходимая для синтеза белков. Мет-тРНКмет участвует в инициации процесса трансляции каждого белка. Как и многие другие аминокислоты, метионин подвергается транс- и дезаминированию. Особая роль метионина заключается в том, что метильная группа этой аминокислоты используется для синтеза целого ряда соединений в реакциях трансметилирования. Основным донором метильной группы является S-аденозилметионин (SAM) - активная форма метионина, который присутствует во всех типах клеток и синтезируется из метионина и АТФ под действием фермента метионин-аденозилтрансферазы:
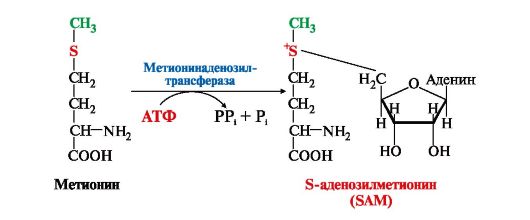 Структура -S+-CH3 в SAM является нестабильной, метильная группа легко отщепляется, что определяет высокую способность ее к переносу на другие соединения в реакциях трансметилирования (рис. 9.20). Структура -S+-CH3 в SAM является нестабильной, метильная группа легко отщепляется, что определяет высокую способность ее к переносу на другие соединения в реакциях трансметилирования (рис. 9.20).
 Рис. 9.20. Метаболизм метионина Рис. 9.20. Метаболизм метионина
В реакциях трансметилирования SAM превращается в S-аденозилгомо- цистеин (SAr), который гидролитически расщепляется с образованием аденозина и гомоцистеина. Последний может снова превращаться в метионин с участием метил-Н4-фолата и витамина В12. Регенерация метионина тесно связана с обменом серина и глицина и взаимопревращениями производных Н4-фолата (см. рис. 9.20).
2. Метионин и серин необходимы для синтеза условно заменимой аминокислоты цистеина, причем в этом процессе метионин является донором атома серы. Цистеин образуется непосредственно из гомоцистеина в ходе двух реакций, которые происходят с участием пиридоксальфосфата (см. рис. 9.20). Генетический дефект этих ферментов приводит к нарушению использования гомоцистеина в организме и превращению его в гомоцистин.
Гомоцистин может накапливаться в крови и тканях, выделяться с мочой, вызывая гомоцистинурию. Заболевание сопровождается эктопией (смещением) хрусталика глаза, катарактой, остеопорозом, умственной отсталостью (-50% больных). Причиной заболевания могут служить как наследственные нарушения обмена гомоцистеина, так и гиповитаминоз фолиевой кислоты или витаминов В12 и
SАМ как донор метильной группы участвует в синтезе многих веществ (лецитина, адреналина, карнитина, ацетилхолина, креатина и др.), а также в инактивации нормальных метаболитов и обезвреживании токсических веществ в печени.
3. Синтез фосфатидилхолина (лецитина) наиболее активно протекает в печени, которая использует лецитин на построение мембран и формирование липопротеинов. Реакцию катализирует фосфатидилэтаноламинтрансметилаза.
 В клетки других тканей фосфатидилхолин доставляется в составе ЛПНП. Особую роль лецитин играет в метаболизме ЛПВП (см. модуль 8). В клетки других тканей фосфатидилхолин доставляется в составе ЛПНП. Особую роль лецитин играет в метаболизме ЛПВП (см. модуль 8).
4. Синтез карнитина - переносчика ацильной группы в митохондрии - происходит путем метилирования γ-аминомасляной кислоты с участием SAM:
 5. Синтез креатина происходит с использованием трех аминокислот: аргинина, глицина и метионина. Процесс начинается в почках, в реакцию вступают аргинин и глицин. Образующийся гуанидинацетат поступает затем в печень, где подвергается метилированию с участием SAM и превращается в креатин. Из печени креатин транспортируется в мышцы и головной мозг. 5. Синтез креатина происходит с использованием трех аминокислот: аргинина, глицина и метионина. Процесс начинается в почках, в реакцию вступают аргинин и глицин. Образующийся гуанидинацетат поступает затем в печень, где подвергается метилированию с участием SAM и превращается в креатин. Из печени креатин транспортируется в мышцы и головной мозг.
Креатин в клетках превращается в креатинфосфат - макроэргическое соединение, являющееся резервной формой энергии в мышечной и нервной тканях. Содержание креатинфосфата в покоящейся мышце в восемь раз выше, чем АТФ. Эту реакцию катализирует фермент креатинкиназа (рис. 9.21).
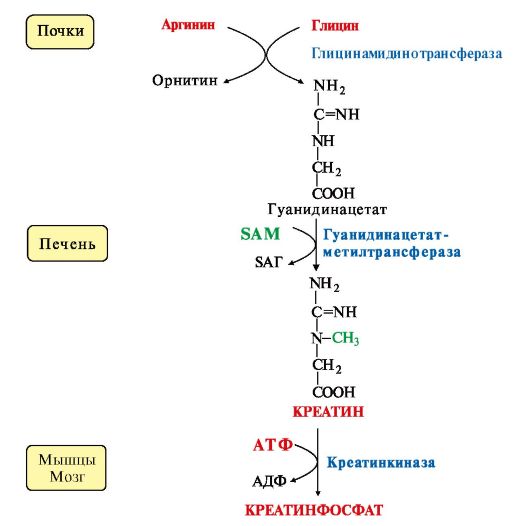 Рис. 9.21. Схема синтеза и использования креатина Рис. 9.21. Схема синтеза и использования креатина
Креатинфосфат играет важную роль в обеспечении работающей мышцы энергией в начальный период физической работы. В работающей мышце концентрация АТФ некоторое время остается постоянной, а концентрация креатинфосфата быстро снижается. Часть образовавшегося креатина и креатинфосфата с постоянной скоростью превращается в креатинин, который выводится с мочой (норма - 1-2 г/сут, или 8,8-17,6 ммоль/л)
При уменьшении массы мышц вследствие длительного отрицательного азотистого баланса, при состояниях, ведущих к атрофии мышц, выделение креатинина снижается (голодание, острые инфекции, сахарный диабет, гипертиреоз и т.д.). Определение содержания в крови креатина и креатинина используется для диагностики заболеваний, а также как показатель эффективности работы мышц в спортивной медицине.
|