Источники атомов пуринового кольца
Содержание книги
- Полиоловый путь превращения глюкозы
- Анаэробный распад глюкозы (гликолиз). Этапы. Гликолитическая оксидоредукция. Регуляция. Физиологическое значение анаэробного распада глюкозы. Баланс энергии.
- Аэробное окисление глюкозы. Изложить анаэробную фазу.
- Изменение скорости реакций цтк и причины накопления кетоновых тел при некоторых состояниях
- Мобилизация гликогена (гликогенолиз)
- Схематичное расположение дефектных ферментов при различных гликогенозах
- Включение глицерина в синтез глюкозы
- Высшие жирные кислоты, структура, свойства, биологическая роль. Понятие о полиненасыщенных жирных кислотах.
- Гликолипиды - рецепторные молекулы
- Липиды пищи, их характеристика. Суточная потребность в липидах.
- Полный ферментативный гидролиз триацилглицерола
- Нарушения переваривания и всасывания пищевых жиров.
- Ресинтез триацилглицеринов в стенке кишечника.
- Формирование кетоновых тел. Причины и последствия кетоза.
- Удлинение цепи жирных кислот
- Реакции синтеза таг из фосфатидной кислоты
- Холестерин, строение, свойства, основные этапы синтеза и его регуляция.
- Пищевые продукты - источники белков. Нормы белка в питании. Биологическая ценность белков. Понятие об азотистом балансе.
- Переваривание белков в желудке, характеристика ферментов. Роль соляной кислоты.
- Изменение кислотности в желудке
- Переваривание белков и полипептидов в кишечнике. Характеристика протеиназ.
- Возможные пути превращений аминокислот
- Схема реакции трансаминирования
- Декарбоксилирование аминокислот. Гистамин, серотонин и другие биогенные амины.
- Появление аммиака в клетках непрерывно
- Реакции глюкозо-аланинового цикла (выделен рамкой).
- Взаимосвязь обмена серина, глицина, метионина и цистеина
- Синтез креатина и креатинина, креатинфосфат - дополнительный источник энергии мышечного сокращения.
- Патология белкового обмена. Белковое голодание. Причины и последствия.
- Врожденные нарушения обмена некоторых аминокислот (фенилкетонурия, алкаптонурия, цистиноз и цистинурия).
- Распад нуклеиновых кислот в тканях. Катаболизм пуриновых оснований.
- Реакции катаболизма пуриновых нуклеотидов
- Источники атомов пуринового кольца
- Биосинтез пиримидиновых нуклеотидов в тканях.
- Нарушения порфиринового обмена. Порфирии.
- Этапы метаболизма билирубина в организме
- Схема патогенеза механической желтухи
- Патологические формы гемоглобина
- Белки сыворотки крови. Отдельные представители. Биологические функции.
- Гамма-глобулины - защитные белки
- Единицы измерения активности ферментов
- Роль почек в регуляции водно-солевого обмена. Вазопрессин, альдостерон, ренин-ангиотензиновая система.
- Физико-химические свойства мочи в норме и ее химический состав.
- У мочи есть и другие особенности
- Патологические компоненты мочи. Клиническое значение анализа мочи.
- Эндогенная вода. Компартментализация жидкостей в организме. Значение воды в организме.
- Биологическая роль, регуляция уровня.
- Биологическая роль, регуляция уровня. Ионы кальция необходимы для передачи нервного импульса
- Регуляция синтеза и секреции. Активируют: Гипокальциемия повышает гидроксилирование витамина D по С1 в почках через
- Железо, его концентрация в сыворотке крови. Биологическая роль. Определение железа в сыворотке крови.
Синтез аденозинмонофосфата и гуанозинмонофосфата
I. Гуанозинмонофосфат (ГМФ) образуется в двух реакциях – сначала ИМФ окисляется ИМФ-дегидрогеназой до ксантозилмонофосфата, источником кислорода является вода, акцептором водорода – НАД. После этого работает ГМФ-синтетаза, она использует универсальный клеточный донор NH2-групп – глутамин, источником энергии для реакции служит АТФ.
II. Аденозинмонофосфат (АМФ) также образуется в двух реакциях, но в качестве донора NH2-группы выступает аспарагиновая кислота. В первой, аденилосукцинат-синтетазной, реакции на присоединение аспартата используется энергия распада ГТФ, во второй реакции аденилосукцинат-лиаза производит удаление части аспарагиновой кислоты в виде фумарата.
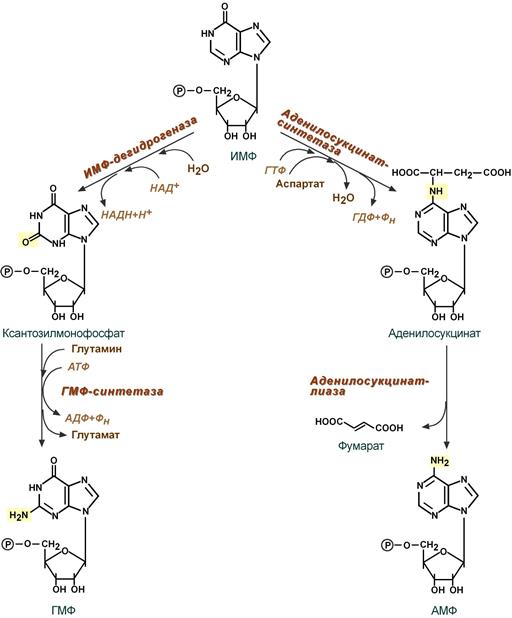
Реакции синтеза АМФ и ГМФ
Образование нуклеозидтрифосфатов АТФ и ГТФ.
Синтез ГТФ осуществляется в 2 стадии посредством переноса макроэргических фосфатных групп от АТФ. Синтез АТФ происходит несколько иначе. АДФ из АМФ образуется также за счет макроэргических связей АТФ. Для синтеза же АТФ из АДФ в митохондриях есть фермент АТФ-синтаза, образующий АТФ в реакциях окислительного фосфорилирования.
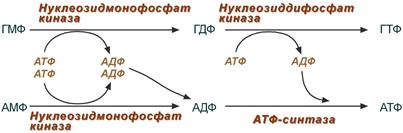
Реакции синтеза АТФ и ГТФ
Синтез пуринов регулируется конечными продуктами
Внутриклеточная регуляция
Регуляция синтеза пуринов (пуриновых нуклеотидов) происходит по механизму обратной отрицательной связи, т.е. продукт реакции (или совокупности реакций) ингибирует начальные этапы процесса. Для синтеза пуринов такими ингибиторами являются АМФ и ГМФ:
· ГМФ блокирует первые две реакции синтеза ИМФ, а также ИМФ-дегидрогеназную реакцию,
· АМФ блокирует первую реакцию синтеза ИМФ и аденилосукцинатсинтетазную реакцию.
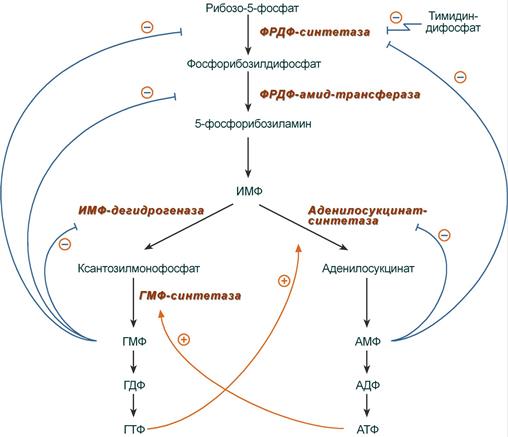
Схема регуляции синтеза пуриновых нуклеотидов
Кроме этого, имеется положительная перекрестная регуляция со стороны АТФ и ГТФ, а именно – как участник реакций каждый из них оказывает стимулирующее влияние на синтез другого нуклеотида. Это заключается в том, что АТФ, принимая участие в ГМФ-синтетазной реакции, облегчает синтез ГМФ. В свою очередь, ГТФ является донором энергии для синтеза АМФ, участвуя в аденилосукцинат-синтетазной реакции.
Пиримидиновый нуклеотид тимидиндифосфат (ТДФ) также блокирует фермент ФРДФ-синтетазу, что снижает синтез ФРДФ, который используется при синтезе не только пуринов, но и пиримидинов.
Лекарственная регуляция
При разработке новых противоопухолевых средств были предложены ингибиторы ферментов:
· ФРДФ-амидтрансферазы – азосерин, диазонорлейцин,
· ИМФ-дегидрогеназы – микофеноловая кислота, 6-меркаптопурин,
· аденилосукцинат-лиазы – 6-меркаптопурин.
|