Возможные пути превращений аминокислот
Содержание книги
- Понятие о метаболических путях. Общие и специфические пути катаболизма углеводов, жиров, аминокислот.
- Роль оксалоацетата в метаболизме
- Строение некоторых производных моносахаридов
- Гетерополисахариды. Отдельные представители. Биологическая роль.
- Полиоловый путь превращения глюкозы
- Анаэробный распад глюкозы (гликолиз). Этапы. Гликолитическая оксидоредукция. Регуляция. Физиологическое значение анаэробного распада глюкозы. Баланс энергии.
- Аэробное окисление глюкозы. Изложить анаэробную фазу.
- Изменение скорости реакций цтк и причины накопления кетоновых тел при некоторых состояниях
- Мобилизация гликогена (гликогенолиз)
- Схематичное расположение дефектных ферментов при различных гликогенозах
- Включение глицерина в синтез глюкозы
- Высшие жирные кислоты, структура, свойства, биологическая роль. Понятие о полиненасыщенных жирных кислотах.
- Гликолипиды - рецепторные молекулы
- Липиды пищи, их характеристика. Суточная потребность в липидах.
- Полный ферментативный гидролиз триацилглицерола
- Нарушения переваривания и всасывания пищевых жиров.
- Ресинтез триацилглицеринов в стенке кишечника.
- Формирование кетоновых тел. Причины и последствия кетоза.
- Удлинение цепи жирных кислот
- Реакции синтеза таг из фосфатидной кислоты
- Холестерин, строение, свойства, основные этапы синтеза и его регуляция.
- Пищевые продукты - источники белков. Нормы белка в питании. Биологическая ценность белков. Понятие об азотистом балансе.
- Переваривание белков в желудке, характеристика ферментов. Роль соляной кислоты.
- Изменение кислотности в желудке
- Переваривание белков и полипептидов в кишечнике. Характеристика протеиназ.
- Возможные пути превращений аминокислот
- Схема реакции трансаминирования
- Декарбоксилирование аминокислот. Гистамин, серотонин и другие биогенные амины.
- Появление аммиака в клетках непрерывно
- Реакции глюкозо-аланинового цикла (выделен рамкой).
- Взаимосвязь обмена серина, глицина, метионина и цистеина
- Синтез креатина и креатинина, креатинфосфат - дополнительный источник энергии мышечного сокращения.
- Патология белкового обмена. Белковое голодание. Причины и последствия.
- Врожденные нарушения обмена некоторых аминокислот (фенилкетонурия, алкаптонурия, цистиноз и цистинурия).
- Распад нуклеиновых кислот в тканях. Катаболизм пуриновых оснований.
- Реакции катаболизма пуриновых нуклеотидов
- Источники атомов пуринового кольца
- Биосинтез пиримидиновых нуклеотидов в тканях.
- Нарушения порфиринового обмена. Порфирии.
- Этапы метаболизма билирубина в организме
- Схема патогенеза механической желтухи
- Патологические формы гемоглобина
- Белки сыворотки крови. Отдельные представители. Биологические функции.
- Гамма-глобулины - защитные белки
- Единицы измерения активности ферментов
- Роль почек в регуляции водно-солевого обмена. Вазопрессин, альдостерон, ренин-ангиотензиновая система.
- Физико-химические свойства мочи в норме и ее химический состав.
- У мочи есть и другие особенности
- Патологические компоненты мочи. Клиническое значение анализа мочи.
- Эндогенная вода. Компартментализация жидкостей в организме. Значение воды в организме.
Реакции превращения аминокислот в клетке условно можно разделить на три части, в зависимости от реагирующей группы:
· с участием аминогруппы - здесь подразумевается удаление от аминокислоты аминогруппы тем или иным способом, в результате чего остается углеводородный скелет,
· по боковой цепи (радикалу) - происходит использование углеродного скелета для синтеза глюкозы, жиров, или для образованеия энергии АТФ,
· по карбоксильной группе - связано с отщеплением карбоксильной группы.
Трансаминирование – реакция переноса альфа-аминогруппы с аминокислоты на альфа-кетокислоту с образованием новой кетокислоты и новой аминокислоты (константа равновесия = 1,0). Процесс легко обратим.

Реакции катализируют ферменты АМИНОТРАНСФЕРАЗЫ, компонентом которых служит пиридоксальфосфат – производн. Витамина В6 (пиридоксина)
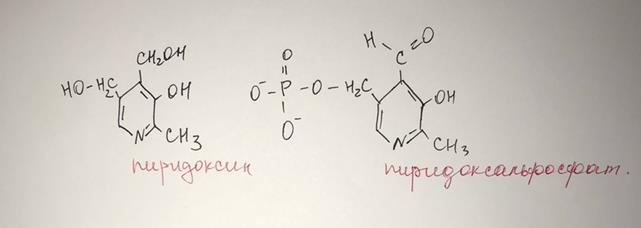
АМТ обнаружены в цитоплазме и в митохондриях.
В реакции трансаминирования не вступают: лизин, треонин, пролин
Весь перенос аминогруппы совершается в две стадии:
· к пиридоксальфосфату сначала присоединяется первая аминокислота, отдает аминогруппу, превращается в кетокислоту и отделяется. Аминогруппа при этом переходит на кофермент и образуется пиридоксаминфосфат.
· на второй стадии к пиридоксаминфосфату присоединяется другая кетокислота, получает аминогруппу, образуется новая аминокислота и пиридоксальфосфат регенерирует.
Реакции трансаминирования:
· активируются в печени, мышцах и других органах при поступлении в клетку избыточного количества тех или иных аминокислот – с целью оптимизации их соотношения,
· обеспечивают синтез заменимых аминокислот в клетке при наличии их углеродного скелета (кетоаналога),
· начинаются при прекращении использования аминокислот на синтез азотсодержащих соединений (белков, креатина, фосфолипидов, пуриновых и пиримидиновых оснований) – с целью дальнейшего катаболизма их безазотистого остатка и выработки энергии,
· необходимы при внутриклеточном голодании, например, при гипогликемиях различного генеза – для использования безазотистого остатка аминокислот в печени для кетогенеза и глюконеогенеза, в других органах – для его прямого вовлечения в реакции цикла трикарбоновых кислот.
При патологиях (сахарный диабет 1 типа, гиперкортицизм) реакции трансаминирования обусловливают появление субстратов для глюконеогенеза и способствуют патологической гипергликемии.
Продукт трансаминирования глутаминовая кислота:
· является одной из транспортных форм аминного азота в гепатоциты,
· способна реагировать со свободным аммиаком, обезвреживая его (синтез глутамина).
Окислительное дезаминирование. Другие виды дезаминирования. Биологическое значение этого процесса.
У человека основным способом дезаминирования является окислительное дезаминирование. Выделяют два варианта окислительного дезаминирования: прямое и непрямое.
|






