Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Формирование кетоновых тел. Причины и последствия кетоза.Содержание книги
Поиск на нашем сайте При состояниях, сопровождающихся снижением глюкозы крови, клетки органов и тканей испытывают энергетический голод. Так как окисление жирных кислот процесс "трудоемкий", а нервные клетки вообще неспособны окислять жирные кислоты, то печень облегчает использование этих кислот тканями, заранее окисляя их до уксусной кислоты и переводя последнюю в транспортную форму – кетоновые тела. К кетоновым телам относят три соединения близкой структуры – ацетоацетат, 3-гидроксибутират и ацетон.
Строение кетоновых тел Стимулом для образования кетоновых тел служит поступление большого количества жирных кислот в печень. Как уже указывалось, при состояниях, активирующих липолиз в жировой ткани, не менее 30% образованных жирных кислот задерживаются печенью. К таким состояниям относится голодание, сахарный диабет I типа, длительные физические нагрузки. Так как синтез ТАГ в этих условиях невозможен, то жирные кислоты из цитозоля попадают в митохондрии и окисляются с образованием кетоновых тел. Для понимания причин развития кетонемии при различных ситуациях необходимо понимать роль оксалоацетата для регуляции цикла трикарбоновых кислот. При голодании соотношение инсулин/глюкагон низкое и поэтому в печени активно идет глюконеогенез, для которого используется оксалоацетат. Поэтому при голодании жирные кислоты, поступающие в большом количестве в печень, не сгорают в ЦТК, а уходят в синтез кетоновых тел. При длительной мышечной нагрузке резервы гликогена в печени истощаются, развивается тенденция к гипогликемии и секретируется глюкагон, который усиливает глюконеогенез, при этом количество оксалоацетата снижается, и жирные кислоты, попавшие в печень, будут окисляться с образованием кетоновых тел. Кроме отмеченных ситуаций, количество кетоновых тел в крови возрастает при алкогольном отравлении и потреблении жирной пищи. При богатой жирами диете, особенно у детей, жирные кислоты не успевают включиться в состав ТАГ и ЛПОНП и частично переходят в митохондрии, что увеличивает синтез кетоновых тел. При алкогольном отравлении субстратом для синтеза кетонов является ацетил-SКоА, синтезируемый при обезвреживании этанола. В обычных условиях синтез кетоновых тел также идет, хотя в гораздо меньшем количестве. Для этого используются как жирные кислоты, так и безазотистые остатки кетогенных и смешанныхаминокислот.
Наиболее частой причиной развития кетоза, а впоследствии – и кетонурии, является сахарный диабет. При сахарном диабете 1-го типа, с одной стороны, имеет место дефицит инсулина, с другой – избыток контринсулярных гормонов (глюкагона, катехоламинов, кортизола). В условиях недостатка инсулина активируются процессы гликолиза, гликогенолиза, липолиза. Массивный липолиз приводит к быстрому увеличению концентрации свободных жирных кислот в крови, из которых в печени под действием глюкагона синтезируются кетоновые кислоты Биосинтез высших жирных кислот. Роль ацетил-КоА в биосинтезе высших жирных кислот, транспорт через митохондриальную мембрану. Биосинтез жирных кислот можно рассматривать как процесс, складывающийся из 3 этапов.
I. Транспорт ацетил-КоА в цитозоль из митохондрий II. Образование малонилКоА III. Конденсация этих молекул и их восстановление с образовани-ем высших насыщенных жирных кислот, главным образом пальмитино-вой.
I этап. Образование ацетил-КоА происходит в митохондриях,а ихмембрана непроницаема для ацетил-КоА. Перенос ацетильных групп происходит при помощи цитрата (цитратный челночный механизм).
Митохондрии
ацетил-КоА + ЩУК
Цитратсинтаза
цитрат + HSKoA
Транслоказа
цитозоль цитрат + АТФ + HSKoA
Цитратлиаза
ацетил-КоА + АДФ + Рн + ЩУК
ЩУК может вернуться в митохондрии с помощью своей транслоказы, но чаще она восстанавливается до малата с участием малатдегид-рогеназы (МДГ).
цитозоль ЩУК+НАДН+Н +
Малат декарбоксилируется НАДФ-зависимой малатдегидро- геназой (маликфермент): МДГ Малат + НАДФ
Образующийся НАДФН2 используется в дальнейшем для синтеза жирных кислот.
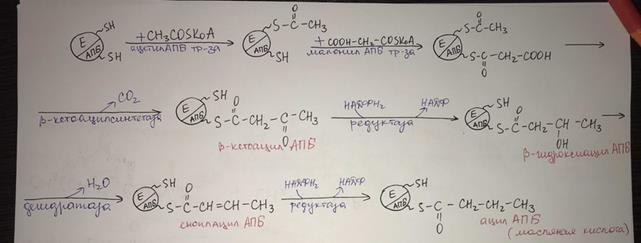
Биосинтез жирных кислот. Общие положения. Формирование малонил-КоА. АПБ, биологическая роль. Стадии удлинения цепочки жирной кислоты. II этап. Ацетил-КоА карбоксилируется под действием ацетил-КоА-карбоксилазы, сложного фермента, коферментом которого служит витамин биотин.
Эта реакция лимитирует скорость всего процесса синтеза жирных кислот.
III этап протекает при участии мультиферментного пальмитат- синтазного комплекса. Он состоит из двух полипептидных цепей.Каждая полипептидная цепь содержит все 6 ферментов синтеза (трансацилаза, кетоацилсинтаза, кетоацилредуктаза, гидратаза, еноилредуктаза, тиоэстераза). Ферменты связаны между собой ковалентными связями, ацилпереносящий белок (АПБ) является также частью полипептидной цепи, но его функция связана только с переносом ацильных радикалов. В процессе синтеза важную роль играют тиогруппы. Одна из них принадлежит 4-фосфопантотеину, входящему в состав АПБ (центральная)
и вторая – цистеину кетоацилсинтазы (периферическая). Функциональная единица синтеза состоит из половины одного мономера, взаимодействующего с комплементарной половиной второго мономера, где центральная SH-группа одного мономера очень близка к периферической SH-группе другого. и Т.е. на синтазном комплексе синтезируются одновременно 2 жирные кислоты и только димер активен. Перенос субстрата от фермента к ферменту происходит при участии АПБ.
|
||
|
|
Последнее изменение этой страницы: 2021-07-18; просмотров: 154; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.217.86 (0.009 с.) |







 МДГ
МДГ малат + НАД +
малат + НАД + пируват + СО 2 + НАДФН 2
пируват + СО 2 + НАДФН 2