Распад нуклеиновых кислот в тканях. Катаболизм пуриновых оснований.
Содержание книги
- Полиоловый путь превращения глюкозы
- Анаэробный распад глюкозы (гликолиз). Этапы. Гликолитическая оксидоредукция. Регуляция. Физиологическое значение анаэробного распада глюкозы. Баланс энергии.
- Аэробное окисление глюкозы. Изложить анаэробную фазу.
- Изменение скорости реакций цтк и причины накопления кетоновых тел при некоторых состояниях
- Мобилизация гликогена (гликогенолиз)
- Схематичное расположение дефектных ферментов при различных гликогенозах
- Включение глицерина в синтез глюкозы
- Высшие жирные кислоты, структура, свойства, биологическая роль. Понятие о полиненасыщенных жирных кислотах.
- Гликолипиды - рецепторные молекулы
- Липиды пищи, их характеристика. Суточная потребность в липидах.
- Полный ферментативный гидролиз триацилглицерола
- Нарушения переваривания и всасывания пищевых жиров.
- Ресинтез триацилглицеринов в стенке кишечника.
- Формирование кетоновых тел. Причины и последствия кетоза.
- Удлинение цепи жирных кислот
- Реакции синтеза таг из фосфатидной кислоты
- Холестерин, строение, свойства, основные этапы синтеза и его регуляция.
- Пищевые продукты - источники белков. Нормы белка в питании. Биологическая ценность белков. Понятие об азотистом балансе.
- Переваривание белков в желудке, характеристика ферментов. Роль соляной кислоты.
- Изменение кислотности в желудке
- Переваривание белков и полипептидов в кишечнике. Характеристика протеиназ.
- Возможные пути превращений аминокислот
- Схема реакции трансаминирования
- Декарбоксилирование аминокислот. Гистамин, серотонин и другие биогенные амины.
- Появление аммиака в клетках непрерывно
- Реакции глюкозо-аланинового цикла (выделен рамкой).
- Взаимосвязь обмена серина, глицина, метионина и цистеина
- Синтез креатина и креатинина, креатинфосфат - дополнительный источник энергии мышечного сокращения.
- Патология белкового обмена. Белковое голодание. Причины и последствия.
- Врожденные нарушения обмена некоторых аминокислот (фенилкетонурия, алкаптонурия, цистиноз и цистинурия).
- Распад нуклеиновых кислот в тканях. Катаболизм пуриновых оснований.
- Реакции катаболизма пуриновых нуклеотидов
- Источники атомов пуринового кольца
- Биосинтез пиримидиновых нуклеотидов в тканях.
- Нарушения порфиринового обмена. Порфирии.
- Этапы метаболизма билирубина в организме
- Схема патогенеза механической желтухи
- Патологические формы гемоглобина
- Белки сыворотки крови. Отдельные представители. Биологические функции.
- Гамма-глобулины - защитные белки
- Единицы измерения активности ферментов
- Роль почек в регуляции водно-солевого обмена. Вазопрессин, альдостерон, ренин-ангиотензиновая система.
- Физико-химические свойства мочи в норме и ее химический состав.
- У мочи есть и другие особенности
- Патологические компоненты мочи. Клиническое значение анализа мочи.
- Эндогенная вода. Компартментализация жидкостей в организме. Значение воды в организме.
- Биологическая роль, регуляция уровня.
- Биологическая роль, регуляция уровня. Ионы кальция необходимы для передачи нервного импульса
- Регуляция синтеза и секреции. Активируют: Гипокальциемия повышает гидроксилирование витамина D по С1 в почках через
- Железо, его концентрация в сыворотке крови. Биологическая роль. Определение железа в сыворотке крови.
В клетках тканей нуклеиновые кислоты распадаются под влиянием нуклеаз. Различают эндонуклеазы и экзонуклеазы. Эндонуклеазы гидролизуют фосфодиэфирные связи одновременно во многих участках молекулы нуклеиновых кислот. По специфичности действия различают ДНКазы и РНКазы. В результате их действия образуются олигонуклеотиды и лишь небольшое количества мононуклеотидов. Экзонуклеазы отщепляют нуклеотиды с 3′ или 5' конца, в результате образуются свободные рибонуклеозид или дезоксирибонуклеозид – 3`или 5' фосфаты, например:
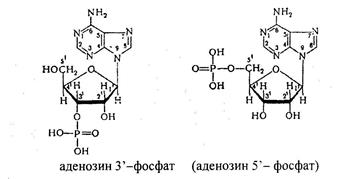
Рибо- и дезоксирибонуклеозидфосфаты расщепляются до нуклеозидов и фосфорной кислоты (под действием фосфатаз).
Нуклеозиды могут расщепляться и гидролитическим путем с участием фермента нуклеозидазы:
Аденозин + Н2О → Аденин + рибоза
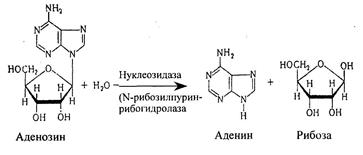
Другие мононуклеотиды подвергаются расщеплению таким же образом.
Пентозы окисляются до CO2 и H2O; фосфорная кислота используется для фосфорилирования органических соединений или выводится из организма. Азотистые основания превращаются в конечные продукты обмена и выделяются с мочой.
Пурины вначале дезаминируются до гипоксантина и ксантина, затем окисляются до конечных продуктов обмена – мочевой кислоты у человека, обезьян, свиней, птиц и аллантоина - у коров, лошадей, собак и кроликов. Мочевая кислота может образовываться из свободных пуринов – аденина и гуанина, а также из соответствующих нуклеозидов и нуклеотидов. Мочевая кислота в печени окисляется до аллантоина:

Гуанин под действием фермента гуаназы превращается в ксантин. Мочевая кислота плохо растворяется в воде (1:15000) и при нарушении обмена нуклеиновых кислот, особенно при избыточном поступлении в организм пуринов, мочевая кислота и ее соли откладываются в виде кристаллов в суставах пальцев, хрящах, в коже и мышцах, образуя узелки. Вокруг узелков развивается очень болезненный воспалительный очаг. Это заболевание известно под названием подагра. У многих животных, кроме млекопитающих, аллантоин может подвергаться дальнейшему расщеплению (у некоторых рыб, амфибий, морских беспозвоночных) с образованиекм аллантоиновой кислоты, а из нее – мочевины и глиоксалевой кислоты.

Пурины распадаются с образованием мочевой кислоты
Наиболее активно катаболизм пуринов идет в печени, тонком кишечнике (пищевые пурины) и почках.
Реакции катаболизма пуринов
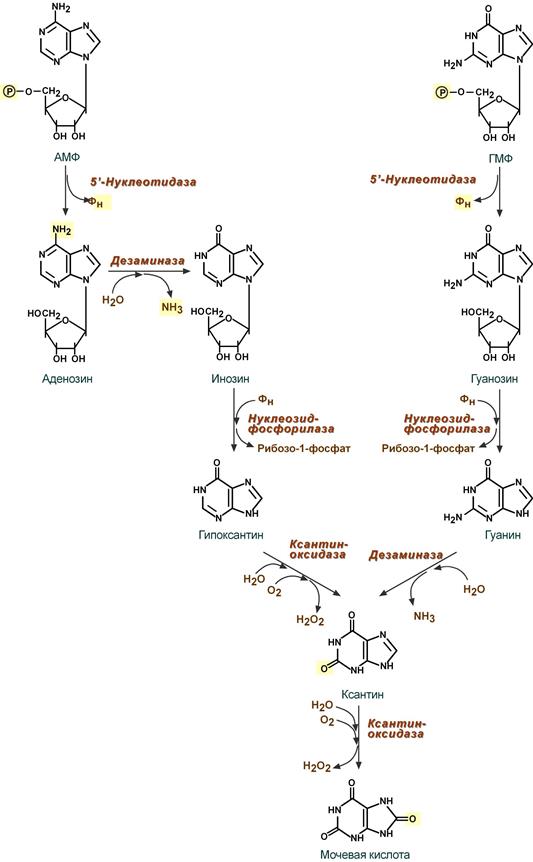
Реакции распада пуринов можно условно разделить на 5 стадий:
1. Дефосфорилирование АМФ и ГМФ – фермент 5'-нуклеотидаза.
2. Гидролитическое отщепление аминогрупы от С6 в аденозине – фермент дезаминаза. Образуется инозин.
3. Удаление рибозы от инозина (с образованием гипоксантина) и гуанозина (с образованием гуанина) с ее одновременным фосфорилированием – фермент нуклеозидфосфорилаза.
4. Окисление С2 пуринового кольца: гипоксантин при этом окисляется до ксантина (фермент ксантиноксидаза), гуанин дезаминируется до ксантина – фермент дезаминаза.
5. Окисление С8 в ксантине с образованием мочевой кислоты – фермент ксантиноксидаза. Около 20% мочевой кислоты удаляется с желчью через кишечник, где она разрушается микрофлорой до CO2 и воды. Остальная часть удаляется через почки.
|