

Мы поможем в написании ваших работ!
ЗНАЕТЕ ЛИ ВЫ?
|
Синтез креатина и креатинина, креатинфосфат - дополнительный источник энергии мышечного сокращения.
Содержание книги
- Полиоловый путь превращения глюкозы
- Анаэробный распад глюкозы (гликолиз). Этапы. Гликолитическая оксидоредукция. Регуляция. Физиологическое значение анаэробного распада глюкозы. Баланс энергии.
- Аэробное окисление глюкозы. Изложить анаэробную фазу.
- Изменение скорости реакций цтк и причины накопления кетоновых тел при некоторых состояниях
- Мобилизация гликогена (гликогенолиз)
- Схематичное расположение дефектных ферментов при различных гликогенозах
- Включение глицерина в синтез глюкозы
- Высшие жирные кислоты, структура, свойства, биологическая роль. Понятие о полиненасыщенных жирных кислотах.
- Гликолипиды - рецепторные молекулы
- Липиды пищи, их характеристика. Суточная потребность в липидах.
- Полный ферментативный гидролиз триацилглицерола
- Нарушения переваривания и всасывания пищевых жиров.
- Ресинтез триацилглицеринов в стенке кишечника.
- Формирование кетоновых тел. Причины и последствия кетоза.
- Удлинение цепи жирных кислот
- Реакции синтеза таг из фосфатидной кислоты
- Холестерин, строение, свойства, основные этапы синтеза и его регуляция.
- Пищевые продукты - источники белков. Нормы белка в питании. Биологическая ценность белков. Понятие об азотистом балансе.
- Переваривание белков в желудке, характеристика ферментов. Роль соляной кислоты.
- Изменение кислотности в желудке
- Переваривание белков и полипептидов в кишечнике. Характеристика протеиназ.
- Возможные пути превращений аминокислот
- Схема реакции трансаминирования
- Декарбоксилирование аминокислот. Гистамин, серотонин и другие биогенные амины.
- Появление аммиака в клетках непрерывно
- Реакции глюкозо-аланинового цикла (выделен рамкой).
- Взаимосвязь обмена серина, глицина, метионина и цистеина
- Синтез креатина и креатинина, креатинфосфат - дополнительный источник энергии мышечного сокращения.
- Патология белкового обмена. Белковое голодание. Причины и последствия.
- Врожденные нарушения обмена некоторых аминокислот (фенилкетонурия, алкаптонурия, цистиноз и цистинурия).
- Распад нуклеиновых кислот в тканях. Катаболизм пуриновых оснований.
- Реакции катаболизма пуриновых нуклеотидов
- Источники атомов пуринового кольца
- Биосинтез пиримидиновых нуклеотидов в тканях.
- Нарушения порфиринового обмена. Порфирии.
- Этапы метаболизма билирубина в организме
- Схема патогенеза механической желтухи
- Патологические формы гемоглобина
- Белки сыворотки крови. Отдельные представители. Биологические функции.
- Гамма-глобулины - защитные белки
- Единицы измерения активности ферментов
- Роль почек в регуляции водно-солевого обмена. Вазопрессин, альдостерон, ренин-ангиотензиновая система.
- Физико-химические свойства мочи в норме и ее химический состав.
- У мочи есть и другие особенности
- Патологические компоненты мочи. Клиническое значение анализа мочи.
- Эндогенная вода. Компартментализация жидкостей в организме. Значение воды в организме.
- Биологическая роль, регуляция уровня.
- Биологическая роль, регуляция уровня. Ионы кальция необходимы для передачи нервного импульса
- Регуляция синтеза и секреции. Активируют: Гипокальциемия повышает гидроксилирование витамина D по С1 в почках через
- Железо, его концентрация в сыворотке крови. Биологическая роль. Определение железа в сыворотке крови.
Креатин – вещество скелетных мышц, миокарда, нервной ткани. В виде креатинфосфата креатин является "депо" макроэргических связей, используется для быстрого ресинтеза АТФ во время работы клетки.
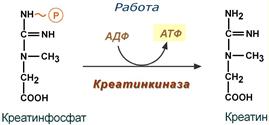
Использование креатинфосфата для ресинтеза АТФ
Особенно показательна роль креатина в мышечной ткани. Креатинфосфат обеспечивает срочный ресинтез АТФ в первые секунды работы (5‑10 сек), когда никакие другие источники энергии (анаэробный гликолиз, аэробное окисление глюкозы, β-окисление жирных кислот) еще не активированы, и кровоснабжение мышцы не увеличено. В клетках нервной ткани креатинфосфат поддерживает жизнеспособность клеток при отсутствии кислорода.
При мышечной работе ионы Са2+, высвободившиеся из саркоплазматического ретикулума, являются активаторами креатинкиназы. Реакция еще интересна тем, что на ее примере можно наблюдать обратную положительную связь — активацию фермента продуктом реакции креатином. Это позволяет избежать снижения скорости реакции по ходу работы, которое должно было бы произойти по закону действующих масс из-за снижения концентрации креатинфосфата в работающих мышцах.
Около 3% креатинфосфата постоянно в реакции неферментативного дефосфорилирования превращается в креатинин. Количество креатинина, выделяемое здоровым человеком в сутки, всегда почти одинаково и зависит только от объема мышечной массы. Уровень активности креатинкиназы в крови и концентрация креатинина в крови и моче являются ценными диагностическими показателями.
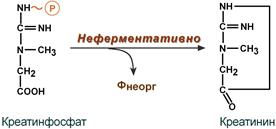
Образование креатинина из креатинфосфата
Синтез креатина
Синтез креатина идет последовательно в почках и печени в двух трансферазных реакциях. По окончании синтеза креатин с током крови доставляется в мышцы или мозг.
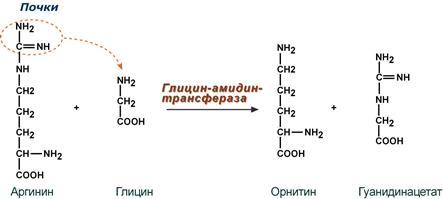

Реакции синтеза креатина в почках и печени
Здесь при наличии энергии АТФ (во время покоя или отдыха) он фосфорилируется с образованием креатинфосфата.
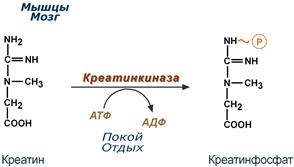
Синтез креатинфосфата
Если синтез креатина опережает возможность его фиксации в мышечной ткани, то развивается креатинурия – появление креатина в моче. Физиологическая креатинурия наблюдается в первые годы жизни ребенка. Иногда к физиологической относят и креатинурию стариков, которая возникает как следствие атрофии мышц и неполного использования образующегося в печени креатина. При заболеваниях мышечной системы (при миопатии или прогрессирующей мышечной дистрофии) в моче наблюдаются наибольшие концентрации креатина – патологическая креатинурия.
|




