Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Формирование пространственных структур и функционирование миоглобина.Содержание книги
Похожие статьи вашей тематики
Поиск на нашем сайте Миоглобин (Мв) - белок, находящийся в красных мышцах, основная функция которого - создание запасов О2, необходимых при интенсивной мышечной работе. Мв - сложный белок, содержащий белковую часть - апоМв и небелковую часть - гем. Первичная структура апоМв определяет его компактную глобулярную конформацию и структуру активного центра, к которому присоединяется небелковая часть миоглобина - гем. Кислород, поступающий из крови в мышцы, связывается с Fe+2 гема в составе миоглобина. Мв - мономерный белок, имеющий очень высокое сродство к О2, поэтому отдача кислорода миоглобином происходит только при интенсивной мышечной работе, когда парциальное давление O2 резко снижается. Формирование конформации Мв. В красных мышцах на рибосомах в ходе трансляции идет синтез первичной структуры Мв, представленной специфической последовательностью 153 аминокислотных остатков. Вторичная структура Мв содержит восемь α-спиралей, называемых латинскими буквами от А до Н, между которыми имеются неспирализованные участки. Третичная структура Мв имеет вид компактной глобулы, в углублении которой между F и Е α-спиралями расположен активный центр (рис. 1.19).
3. Особенности строения и функционирования активного центра Мв. Активный центр Мв сформирован преимущественно гидрофобными радикалами аминокислот, далеко отстоящими друг от друга в первичной структуре (например, Три 39 и Фен138) К активному центру присоединяется плохо растворимые в воде лиганды - гем и О2. Гем - специфический лиганд апоМв (рис. 1.20), основу которого составляют четыре пиррольных кольца, соединенных метенильными мостиками; в центре расположен атом Fe+2, соединенный с атомами азота пиррольных колец четырьмя координационными связями. В активном центре Мв кроме гидрофобных радикалов аминокислот имеются также остатки двух аминокислот с гидрофильными радикалами - Гис Е7 (Гис64) и Гис F8 (Гис93) (рис. 1.21).
Гем через атом железа ковалентно связан с Гис F8. O2 присоединяется к железу с другой стороны плоскости гема. Гис Е7необходим для правильной ориентации О2 и облегчает присоединение кислорода к Fe+2 гема Гис F8 образует координационную связь с Fe+2 и прочно фиксирует гем в активном центре. Гис Е7 необходим для правильной ориентации в активном центре другого лиганда - O2 при его взаимодействии с Fe+2 гема. Микроокружение гема создает условия для прочного, но обратимого связывания O2 с Fe+2 и препятствует попаданию в гидрофобный активный центр воды, что может привести к его окислению в Fе+3. Мономерное строение Мв и его активного центра определяет высокое сродство белка к О2. 4. Олигомерное строение Нв и регуляция сродства Нв к О2 лигандами. Гемоглобины человека - семейство белков, так же как и миоглобин относящиеся к сложным белкам (гемопротеинам). Они имеют тетрамерное строение и содержат две α-цепи, но различаются по строению двух других полипептидных цепей (2α-, 2х-цепи). Строение второй полипептидной цепи определяет особенности функционирования этих форм Нв. Около 98% гемоглобина эритроцитов взрослого человека составляет гемоглобин А (2α-, 2р-цепи). В период внутриутробного развития функционируют два основных типа гемоглобинов: эмбриональный Нв (2α, 2ε), который обнаруживается на ранних этапах развития плода, и гемоглобин F (фетальный) - (2α, 2γ), который приходит на смену раннему гемоглобину плода на шестом месяце внутриутробного развития и только после рождения замещается на Нв А. Нв А - белок, родственный миоглобину (Мв), содержится в эритроцитах взрослого человека. Строение его отдельных протомеров аналогично таковому у миоглобина. Вторичная и третичная структуры миоглобина и протомеров гемоглобина очень сходны, несмотря на то что в первичной структуре их полипептидных цепей идентичны только 24 аминокислотных остатка (вторичная структура протомеров гемоглобина, так же как миоглобин, содержит восемь α-спиралей, обозначаемых латинскими буквами от А до Н, а третичная структура имеет вид компактной глобулы). Но в отличие от миоглобина гемоглобин имеет олигомерное строение, состоит из четырех полипептидных цепей, соединенных нековалентными связями (рис 1.22). Каждый протомер Нв связан с небелковой частью - гемом и соседними протомерами. Соединение белковой части Нв с гемом аналогично таковому у миоглобина: в активном центре белка гидрофобные части гема окружены гидрофобными радикалами аминокислот за исключением Гис F8 и Гис Е7, которые расположены по обе стороны от плоскости гема и играют аналогичную роль в функционировании белка и связывании его с кислородом (см. строение миоглобина).
Кроме того, Гис Е7 выполняет важную дополнительную роль в функционировании Нв. Свободный гем имеет в 25 000 раз более высокое сродство к СО, чем к О2. СО в небольших количествах образуется в организме и, учитывая его высокое сродство к гему, он мог бы нарушать транспорт необходимого для жизни клеток О2. Однако в составе гемоглобина сродство гема к оксиду углерода превышает сродство к О2 всего в 200 раз благодаря наличию в активном центре Гис Е7. Остаток этой аминокислоты создает оптимальные условия для связывания гема с O2 и ослабляет взаимодействие гема с СО. 5. Основная функция Нв - транспорт О2 из легких в ткани. В отличие от мономерного миоглобина, имеющего очень высокое сродство к О2 и выполняющего функцию запасания кислорода в красных мышцах, олигомерная структура гемоглобина обеспечивает: 1) быстрое насыщение Нв кислородом в легких; 2) способность Нв отдавать кислород в тканях при относительно высоком парциальном давлении O2 (20-40 мм рт. ст.); 3) возможность регуляции сродства Нв к О2. 6. Кооперативные изменения конформации протомеров гемоглобина ускоряют связывание O2 в легких и отдачу его в ткани. В легких высокое парциальное давление O2 способствует связыванию его с Нв в активном центре четырех протомеров (2α и 2β). Активный центр каждого протомера, так же как и в миоглобине, расположен между двумя α-спиралями (F и Е) в гидрофобном кармане. Он содержит небелковую часть - гем, прикрепленный к белковой части множеством слабых гидрофобных взаимодействий и одной прочной связью между Fe2+ гема и Гис F8 (см. рис. 1.21). В дезоксигемоглобине, благодаря этой связи с Гис F8, атом Fe2+ выступает из плоскости гема по направлению к гистидину. Связывание O2 с Fe2+ происходит по другую сторону гема в области Гис Е7 с помощью единственной свободной координационной связи. Гис Е7 обеспечивает оптимальные условия для связывания O2 с железом гема. Присоединение O2 к атому Fe+2 одного протомера вызывает его перемещение в плоскость гема, а за ним и остатка гистидина, связанного с ним (рис. 1.23).
Это приводит к изменению конформации всех полипептидных цепей за счет их конформационной лабильности. Изменение конформации других цепей облегчает их взаимодействие со следующими молекулами О2. Четвертая молекула О2 присоединяется к гемоглобину в 300 раз легче, чем первая (рис. 1.24).
В тканях каждая следующая молекула O2 отщепляется легче, чем предыдущая, также за счет кооперативных изменений конформации протомеров. 7. CO2 и Н+, образующиеся при катаболизме органических веществ, уменьшают сродство гемоглобина к О2 пропорционально их концентрации. Энергия, необходимая для работы клеток, вырабатывается преимущественно в митохондриях при окислении органических веществ с использованием O2, доставляемого из легких гемоглобином. В результате окисления органических веществ образуются конечные продукты их распада: СО2 и K2O, количество которых пропорционально интенсивности протекающих процессов окисления. СO2 диффузией попадает из клеток в кровь и проникает в эритроциты, где под действием фермента карбангидразы превращается в угольную кислоту. Эта слабая кислота диссоциирует на протон и бикарбонат ион.
Увеличение количества освобожденного кислорода в зависимости от увеличения концентрации Н+ в эритроцитах называется эффектом Бора (по имени датского физиолога Христиана Бора, впервые открывшего этот эффект). В легких высокое парциальное давление кислорода способствует его связыванию с дезоксиНв, что уменьшает сродство белка к Н+. Освободившиеся протоны под действием карбангидразы взаимодействуют с бикарбонатами с образованием СО2 и Н2О
А - влияние концентрации СО2 и Н+ на высвобождение О2 из комплекса с Нв (эффект Бора); Б - оксигенирование дезоксигемоглобина в легких, образование и выделение СО2. Образовавшийся СО2 поступает в альвеолярное пространство и удаляется с выдыхаемым воздухом. Таким образом, количество высвобождаемого гемоглобином кислорода в тканях регулируется продуктами катаболизма органических веществ: чем интенсивнее распад веществ, например при физических нагрузках, тем выше концентрация СО2 и Н+ и тем больше кислорода получают ткани в результате уменьшения сродства Нв к О2. 8. Аллостерическая регуляция сродства Нв к О2 лигандом - 2,3-бис- фосфоглицератом. В эритроцитах из продукта окисления глюкозы - 1,3-бисфосфоглицерата синтезируется аллостерический лиганд гемоглобина - 2,3-бисфосфоглицерат (2,3-БФГ). В нормальных условиях концентрация 2,3-БФГ высокая и сравнима с концентрацией Нв. 2,3-БФГ имеет сильный отрицательный заряд -5.
В центре тетрамерной молекулы гемоглобина находится полость. Ее образуют аминокислотные остатки всех четырех протомеров (см. рис. 1.22). В капиллярах тканей протонирование Нв (эффект Бора) приводит к разрыву связи между железом гема и О2. В молекуле дезоксигемоглобина по сравнению с оксигемоглобином возникают дополнительные ионные связи, соединяющие протомеры, вследствие чего размеры центральной полости по сравнению с оксигемоглобином увеличиваются. Центральная полость является местом присоединения 2,3-БФГ к гемоглобину. Из-за различия в размерах центральной полости 2,3-БФГ может присоединяться только к дезоксигемоглобину. 2,3-БФГ взаимодействует с гемоглобином в участке, удаленном от активных центров белка и относится к аллостерическим (регуляторным) лигандам, а центральная полость Нв является аллостерическим центром. 2,3-БФГ имеет сильный отрицательный заряд и взаимодействует с пятью положительно заряженными группами двух β-цепей Нв: N-концевой α-аминогруппой Вал и радикалами Лиз82 Гис143 (рис. 1.26).
БФГ связывается с тремя положительно заряженными группами в каждой β-цепи. В капиллярах тканей образующийся дезоксигемоглобин взаимодействует с 2,3-БФГ и между положительно заряженными радикалами β-цепей и отрицательно заряженным лигандом образуются ионные связи, которые изменяют конформацию белка и снижают сродство Нв к О2. Уменьшение сродства Нв к О2 способствует более эффективному выходу О2 в ткани. В легких при высоком парциальном давлении кислород взаимодействует с Нв, присоединяясь к железу гема; при этом изменяется конформация белка, уменьшается центральная полость и происходит вытеснение 2,3-БФГ из аллостерического центра
пространственно удаленных друг от друга (аллостерических), способно вызывать конформационные изменения во всей белковой молекуле. Благодаря взаимодействию с регуляторными лигандами происходит изменение конформации и адаптация функции белковой молекулы к изменениям окружающей среды.
|
||||||||
|
|
Последнее изменение этой страницы: 2017-01-24; просмотров: 1464; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.60 (0.011 с.) |

 Рис. 1.19. Структура миоглобина
Рис. 1.19. Структура миоглобина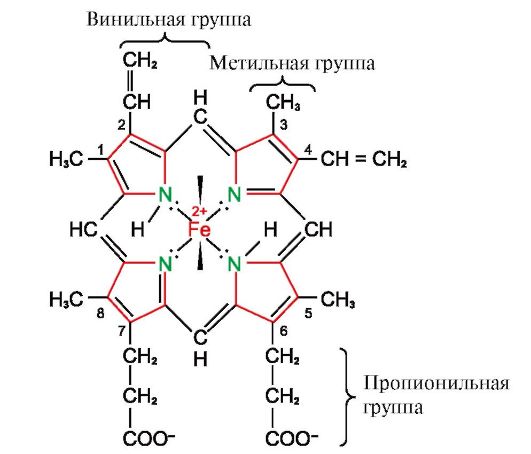 Рис. 1.20. Строение гема - небелковой части миоглобина и гемоглобина
Рис. 1.20. Строение гема - небелковой части миоглобина и гемоглобина Рис. 1.21. Расположение гема и O2 в активном центре апомиоглобина и протомеров гемоглобина
Рис. 1.21. Расположение гема и O2 в активном центре апомиоглобина и протомеров гемоглобина Рис. 1.22. Олигомерная структура гемоглобина
Рис. 1.22. Олигомерная структура гемоглобина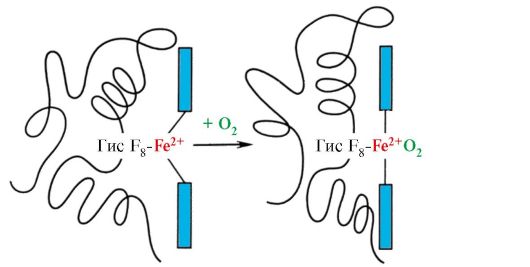 Рис. 1.23. Изменение конформации протомера гемоглобина при соединении с O2
Рис. 1.23. Изменение конформации протомера гемоглобина при соединении с O2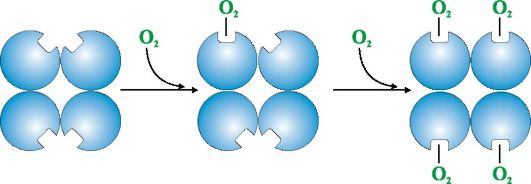 Рис. 1.24. Кооперативные изменения конформации протомеров гемоглобина при его взаимодействии с О2
Рис. 1.24. Кооперативные изменения конформации протомеров гемоглобина при его взаимодействии с О2 Н+ способны присоединятся к радикалам Гис 146 в α- и β-цепях гемоглобина, т.е. в участках, удаленных от гема. Протонирование гемоглобина снижает его сродство к О2, способствует отщеплению О2 от оксиНв, образованию дезоксиНв и увеличивает поступление кислорода в ткани пропорционально количеству образовавшихся протонов (рис. 1.25).
Н+ способны присоединятся к радикалам Гис 146 в α- и β-цепях гемоглобина, т.е. в участках, удаленных от гема. Протонирование гемоглобина снижает его сродство к О2, способствует отщеплению О2 от оксиНв, образованию дезоксиНв и увеличивает поступление кислорода в ткани пропорционально количеству образовавшихся протонов (рис. 1.25).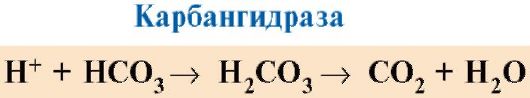
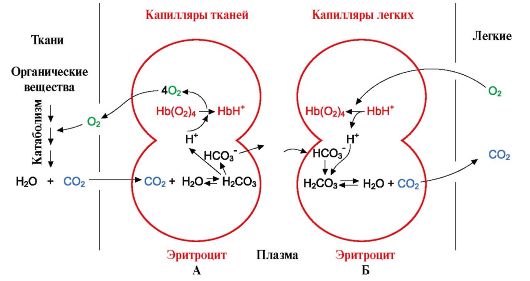 Рис. 1.25. Зависимость сродства Нв к О2 от концентрации СО2 и протонов (эффект Бора):
Рис. 1.25. Зависимость сродства Нв к О2 от концентрации СО2 и протонов (эффект Бора):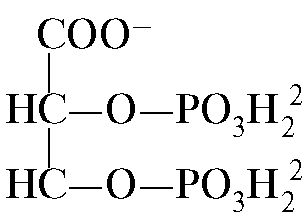 Бисфосфоглицерат в капиллярах тканей, связываясь с дезоксигемоглобином, увеличивает выход кислорода в ткани, уменьшая сродство Нв к О2.
Бисфосфоглицерат в капиллярах тканей, связываясь с дезоксигемоглобином, увеличивает выход кислорода в ткани, уменьшая сродство Нв к О2. Рис. 1.26. БФГ в центральной полости дезоксигемоглобина
Рис. 1.26. БФГ в центральной полости дезоксигемоглобина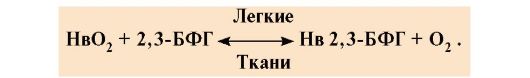 Таким образом, олигомерные белки обладают новыми по сравнению с мономерными белками свойствами. Присоединение лигандов на участках,
Таким образом, олигомерные белки обладают новыми по сравнению с мономерными белками свойствами. Присоединение лигандов на участках,