

Мы поможем в написании ваших работ!
ЗНАЕТЕ ЛИ ВЫ?
|
Обмен нуклеопротеидов: синтез и распад пуринов.
Содержание книги
- Структура и функция аминокислот с полярным (заряженным положительно) радикалом. Роль радикалов гистидина в гемоглобине и ферментах. Роль радикалов лизина в структурах коллагена и эластина. Гистоны.
- Методы идентификации и разделения аминокислот
- Химическая модификация аминокислотных радикалов в белках и их роль в построении структуры и функции белков.
- Структура и функции сложных белков: гликопротеиды, протеогликаны, нуклеопротеиды, фосфопротеиды, липопротеиды, хромопротеиды, металлопротеиды.
- Регуляция активности ферментов: неспецифическая, аллостерическая, регуляция на уровне индукции и репрессии, гормональная регуляция.
- Распределение ферментов в клетках, тканях, органах и биологических жидкостях. Мультиферментные комплексы, преимущества организации
- Наследственные энзимопатии - это заболевания, вызванные наследственными нарушениями биосинтеза ферментов или их структуры и функции.
- Структура, классификация и роль углеводов в организме, пищеварение в жкт у новорождённых и детей более старших возрастов, всасывание продуктов гидролиза
- Биологическая роль углеводов
- Аэробный гликолиз: стадия, ферменты и коферменты, биоэнергетика значение для организма. Челночные механизмы транспорта атома водорода в митохондрии и их роль в поддержании аэробного гликолиза.
- Пути вхождения углеводов в гликолиз. Полное окисление фруктозы (начиная с фруктокиназной реакции). Биоэнергетика процесса, фруктозурия.
- Гликогенез и гликогенолиз в печени. Роль печени в поддержании нормального уровня глюкозы в крови.
- Роль печени в поддержании нормального уровня содержания глюкозы в крови.
- Наследственные патологии углеводного обмена: гликогенозы, фруктозурия, галактоземия, непереносимость лактозы и др.
- Липиды организма: классификация, строение, физико-химические свойства, биологическая роль. Пищеварение липидов в жкт новорожденных
- Энтерогепатический цикл холестерола и желчных кислот.
- Синтез гликогена из глицерола в печени.
- Использование кетоновых тел .
- Роль гормонов в регуляции липидного обмена. Патология липидного обмена (жировая инфильтрация печени, атеросклероз, ожирение, наследственные заболевания), причины возникновения и последствия.
- Мембрана эндоплазматического ретикулума (ЭР)
- Переваривание белков в жкт, биосинтез соляной кислоты и пепсиногена в желудке. Протеазы панкреаса и кишечника. Активирование зимогенов в жкт, специфика действия каждой из протеаз.
- Метаболизм аргинина. Окисление аргинина до конечных продуктов, биоэнергетика процесса. Орнитиновый цикл.
- Интеграция обмена аминокислот с обменом других соединений. Синтез глюкозы из глутамата в печени.
- Метаболизм амидного азота глутамина в почках
- Наследственные патологии обмена фенилаланина, тирозина,лейцина,цистеина. Изменения в белковом обмене при сахарном диабете
- Обмен хромопротеидов: синтез и распад гемма. Обтурационная, паренхиматозная и гемолитическая желтуха. Порфирии.
- Обмен нуклеопротеидов: синтез и распад пуринов.
- Этапы реализации генетической информации: репликация, транскрипция, трансляция.
- Особенности метаболизма лейкоцитов.
- Свертывание крови: внешние и внутренние пути.
- Перенос углекислого газа кровью от тканей к легким
- Гипоксия при патологических процессах, нарушающих снабжение или утилизацию кислорода тканями.
- Нарушения кислотно-основного равновесия
- Нарушения общего объема крови
- Патологические изменения эритроцитов
- Распределение в организме и участие в бихимических реакциях мкро и микроэлементов: наприя,калия, кальция,магния,кобольта,железа,цинка,фосфора,хлора и фтора.
- А. Механизм действия гидрофильных гормонов
- А. Механизм действия липофильных гормонов
- Гормоны щитовидной железы,влияние на обмен в-в. Гипо и гипертиреозы. Механизмы возникновения и последствия.
Нуклеопротеиды представляют собой вещества, построенные из белка и различных нуклеиновых кислот (днк и рнк). ДНК является важнейшим компонентом в хромосомах и имеет тесное отношение к передаче наследственных свойств.РНК хорошо определяется в цитоплазме в виде плотных перинуклеарных скоплений. Нукл.к-ты имеют два источника: поступление с пищей и саморепродукция. Нуклеиновые кислоты состоят из нуклеотидов, которые в свою очередь состоят из: азотистого основания (пуринового или пиримидинового),углевода (рибоза или дезоксирибоза) и фосфорной кислоты. Распад нуклеотидов дает мочевую кислоту, которая как конечный продукт пуринового обмена выделяется с мочой. К патологии нуклеопротеидов относится мочекислый инфаркт и подагара.
Синтез пуринов. Для того, чтобы синтезировать адениловый или гуаниловый нуклеотид, необходимо получить инозиновую кислоту (инозинмонофосфат; IMP). На схеме 35.3 представлена схема пути биосинтеза пуриновых нуклеотидов, а именно IMP. Реакция 1 - образование 5-фосфорибозил-1-пирофосфата (ФРПФ), он необходим для синтеза NAD и NADP — двух коферментов, в состав которых входит никотиновая кислота.присоединяется аминогруппа, которая впоследствии в кольце становится N-9. Глицин и формильная группа N10-формилтетрагидрофолата поставляют недостающие атомы пятичленного кольца. Прежде чем это кольцо замкнется, присоединяются атомы N-3 и N-6 шестичленного кольца. Затем построение кольца продолжается путем присоединения N-1 и С-2. На последней стадии шестичленное кольцо замыкается с образованием инозин-5'-монофосфата [ ИМФ (IMP)], который, однако, не накапливается, а быстро превращается в АМФ и ГМФ.
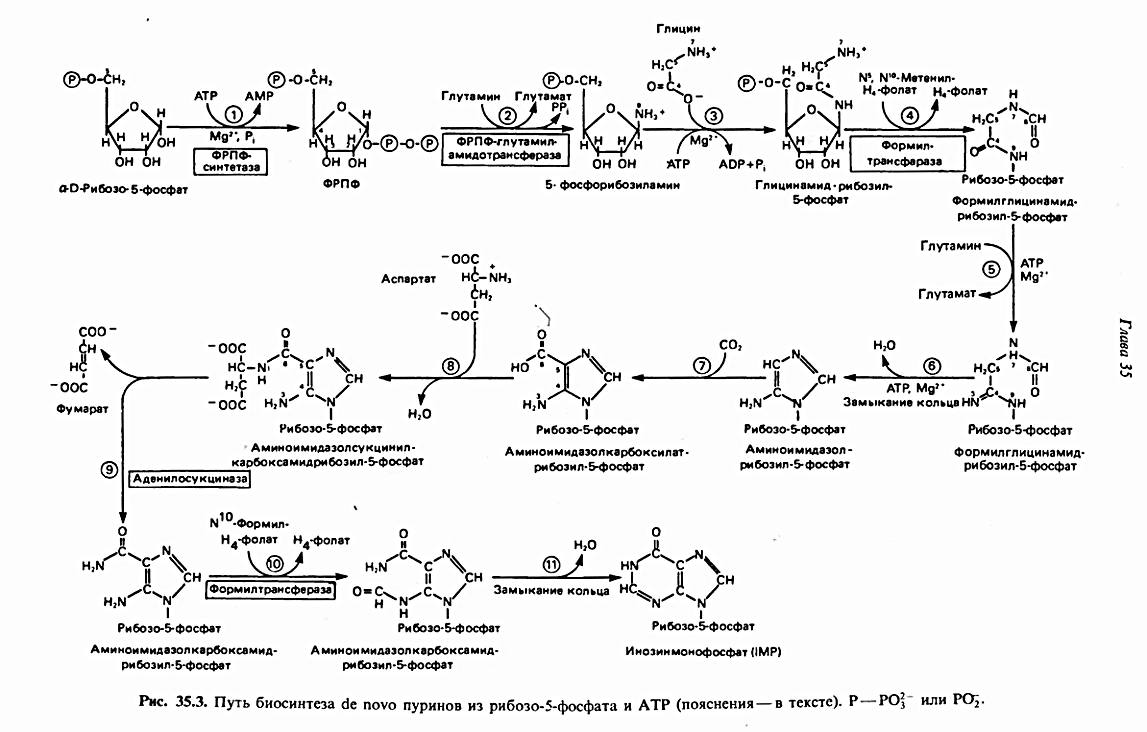
Синтез пуринов (Аденин и Гуанин) объяснение к схеме
Адениновые (реакции 12 и 13) и гуаниновые нуклеотиды (реакции 14 и 15) образуются путем аминирования и соответственно окисления и аминирования общего предшественника—инозинмонофосфата (IMP). Аминирование ГМР протекает через стадию образования промежуточного соединения, в котором аспартат присоединяется к инозиновой кислоте, образуя аденилосукцинат. Отщепление фумарата от аденилосукцината катализируется ферментом аденилосукциназой.
Так же, в две стадии, из IMP образуется гуанзинмонофосфат (GMP). В первой реакции на этом пути (реакция 14) при участии NAD и происходит окисление IMP с образованием ксантинмонофосфата (ХМР). Затем ХМР аминируется амидогруппой глутамина (реакция 15). Для этого процесса необходим АТР, что в какой-то мере напоминает потребность в GTP при превращении IMP в АМР.

Распад пуринов
Конечный продукт катаболизма пуринов у человека — мочевая кислота (образуется преимущественно в печени, выделяется почками как конечный продукт обмена пуриновых нуклеотидов).При этом кольцевая структура пиримидина не разрушается. Установлено, что 99% мочевой кислоты образуется из субстратов нуклеозидфосфорилазы. Пуриновые продукты нуклеозидфосфорилазной реакции — гипоксантин и гуанин — превращаются в мочевую кислоту; промежуточным продуктом является ксантин, образующийся в реакциях, катализируемых гуаназой и ксантинокеидазой в печени, тонком кишечнике и почках. Ксантиноксидаза представляет собой важную мишень для фармакологического вмешательства при гиперурикемии и подагре.
80.Синтез и распад пиримидинов.
Непосредственными предшественниками в синтезе пиримидинового кольца являются карбамоилфосфат, который образуется из глутамина и НСО3-и аспартат. После образования N-карбамоиласпартата происходит замыкание кольца с образованием дигидрооротата. Далее дигидрооротат окисляется флавинмо-нонуклеотидзависимой дегидрогеназой в оротат, который связывается с фосфорибозилдифосфатом (PRPP) с образованием нуклеотида оротидин-5'-монофосфата [ ОМФ (ОМР)], декарбоксилирование которого приводит к уридин-5'-монофосфату [ УМФ (UMP)].
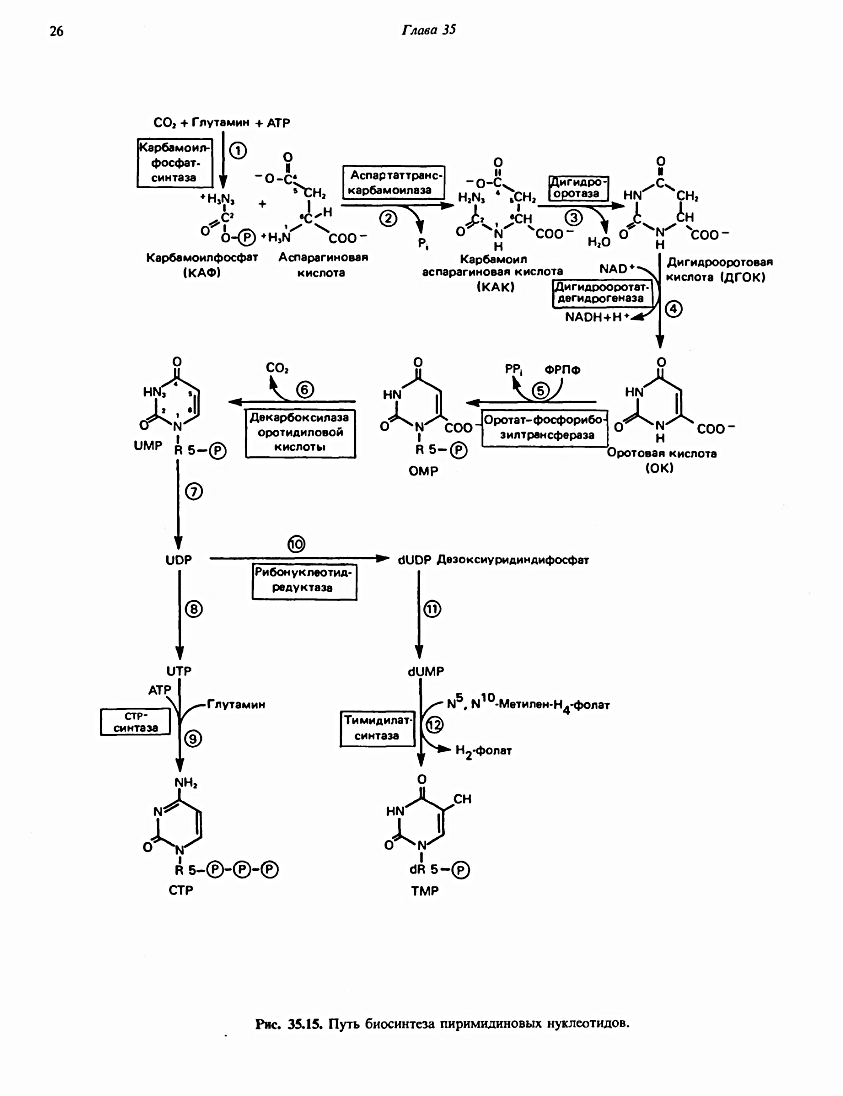
Катаболизм пиримидинов
В отличие от пуринов, кольцевая структура пиримидинов разрушается с образованием обычных конечных продуктов катаболизма - β-аминокислот, аммиака и двуокиси углерода. В катаболизме пиримидиновых нуклеотидов принимают участие нуклеотидазы и пиримидиновые нуклеотид фосфорилазы, которые превращают мононуклеотиды в свободные основания. Аминогруппы цитозина и 5-метилцитозина отделяется в форме аммиака.
|




