

Мы поможем в написании ваших работ!
ЗНАЕТЕ ЛИ ВЫ?
|
Синтез гликогена из глицерола в печени.
Содержание книги
- Структура и функция аминокислот с полярным (заряженным положительно) радикалом. Роль радикалов гистидина в гемоглобине и ферментах. Роль радикалов лизина в структурах коллагена и эластина. Гистоны.
- Методы идентификации и разделения аминокислот
- Химическая модификация аминокислотных радикалов в белках и их роль в построении структуры и функции белков.
- Структура и функции сложных белков: гликопротеиды, протеогликаны, нуклеопротеиды, фосфопротеиды, липопротеиды, хромопротеиды, металлопротеиды.
- Регуляция активности ферментов: неспецифическая, аллостерическая, регуляция на уровне индукции и репрессии, гормональная регуляция.
- Распределение ферментов в клетках, тканях, органах и биологических жидкостях. Мультиферментные комплексы, преимущества организации
- Наследственные энзимопатии - это заболевания, вызванные наследственными нарушениями биосинтеза ферментов или их структуры и функции.
- Структура, классификация и роль углеводов в организме, пищеварение в жкт у новорождённых и детей более старших возрастов, всасывание продуктов гидролиза
- Биологическая роль углеводов
- Аэробный гликолиз: стадия, ферменты и коферменты, биоэнергетика значение для организма. Челночные механизмы транспорта атома водорода в митохондрии и их роль в поддержании аэробного гликолиза.
- Пути вхождения углеводов в гликолиз. Полное окисление фруктозы (начиная с фруктокиназной реакции). Биоэнергетика процесса, фруктозурия.
- Гликогенез и гликогенолиз в печени. Роль печени в поддержании нормального уровня глюкозы в крови.
- Роль печени в поддержании нормального уровня содержания глюкозы в крови.
- Наследственные патологии углеводного обмена: гликогенозы, фруктозурия, галактоземия, непереносимость лактозы и др.
- Липиды организма: классификация, строение, физико-химические свойства, биологическая роль. Пищеварение липидов в жкт новорожденных
- Энтерогепатический цикл холестерола и желчных кислот.
- Синтез гликогена из глицерола в печени.
- Использование кетоновых тел .
- Роль гормонов в регуляции липидного обмена. Патология липидного обмена (жировая инфильтрация печени, атеросклероз, ожирение, наследственные заболевания), причины возникновения и последствия.
- Мембрана эндоплазматического ретикулума (ЭР)
- Переваривание белков в жкт, биосинтез соляной кислоты и пепсиногена в желудке. Протеазы панкреаса и кишечника. Активирование зимогенов в жкт, специфика действия каждой из протеаз.
- Метаболизм аргинина. Окисление аргинина до конечных продуктов, биоэнергетика процесса. Орнитиновый цикл.
- Интеграция обмена аминокислот с обменом других соединений. Синтез глюкозы из глутамата в печени.
- Метаболизм амидного азота глутамина в почках
- Наследственные патологии обмена фенилаланина, тирозина,лейцина,цистеина. Изменения в белковом обмене при сахарном диабете
- Обмен хромопротеидов: синтез и распад гемма. Обтурационная, паренхиматозная и гемолитическая желтуха. Порфирии.
- Обмен нуклеопротеидов: синтез и распад пуринов.
- Этапы реализации генетической информации: репликация, транскрипция, трансляция.
- Особенности метаболизма лейкоцитов.
- Свертывание крови: внешние и внутренние пути.
- Перенос углекислого газа кровью от тканей к легким
- Гипоксия при патологических процессах, нарушающих снабжение или утилизацию кислорода тканями.
- Нарушения кислотно-основного равновесия
- Нарушения общего объема крови
- Патологические изменения эритроцитов
- Распределение в организме и участие в бихимических реакциях мкро и микроэлементов: наприя,калия, кальция,магния,кобольта,железа,цинка,фосфора,хлора и фтора.
- А. Механизм действия гидрофильных гормонов
- А. Механизм действия липофильных гормонов
- Гормоны щитовидной железы,влияние на обмен в-в. Гипо и гипертиреозы. Механизмы возникновения и последствия.
1. глицерин-> глицеролфосфат (глицеролкиназа)
СН2(ОH)-CH2(OH)-CH2(OH) +2AТФ+ Mg2+à СН2(ОH)-CH2(OH)-CH2OPO3+ 2AДФ
2. глицеролфосфат à диоксиацетонфосфат (глицеролфосфатдегидрогеназа)
СН2(ОH)-CH(OH)-CH2OPO3+2НАД+àCH2(OH)-C=O-CH2OPO3 + 2НАДН +Н+
3. диоксиацетонфосфат à глицеральдегид-3-фосфат (триозофосфатизомераза)
CH2(OH)-C=O-CH2OPO3à HC(O)-CH(OH)-CH2OPO3
4. глицеральдегид-3-фосфат à фруктозо-1,6- бисфосфат (глицеральдегидфосфатдегидрогеназа)
 HC(O)-CH(OH)-CH2OPO3à HC(O)-CH(OH)-CH2OPO3à
5. фруктозо -1,6- бисфосфат à фруктозо -6- фосфат
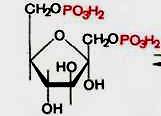 +H2O à +H2O à 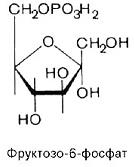
6. фруктозо -6- фосфат à глюкозо -6- фосфат (фосфоглюкоизомераза)
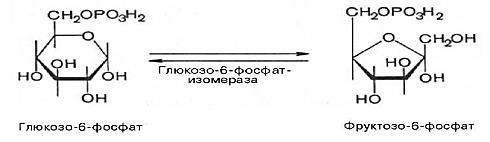
7. глюкозо-6-фосфат à глюкозо-1-фосфат (фосфоглюкомутаза)

8. Глюкозо-1-фосфат + УТФ < = > УДФ-глюкоза + ФнФн (глюкозо-1-фосфатуридилтрансфераза)
.

49. Биосинтез жирных кислот. Локализация, цитратный челнок, образование малонил-КоА, этапы синтеза, источники НАДФН, биоэнергетика процесса.
Биосинтез насыщенных жирных кислот происходит во всех живых организмах в жировой ткани,молочной железе и в печени. Процесс осуществляется в цитоплазме, эндоплазматическом ретикулуме и митохондриях с участием мультиферментного комплекса синтетаза высших жирных кислот. Этот комплекс состоит из шести ферментов, связанных с ацилпереносящим белком, который содержит две свободные SH-группы (АПБ-SH). Синтез происходит путём полимеризации двууглеродных фрагментов, конечным продуктом его является пальмитиновая кислота – насыщенная жирная кислота, содержащая 16 атомов углерода. Обязательными компонентами, участвующими в синтезе, являются НАДФН (кофермент, образующийся в реакциях пентозофосфатного пути окисления углеводов) и АТФ.
Насыщенные жирные кислоты образуются из ацетила-КоА (образуется в митохондриях из пирувата – продукта гликолитического распада глюкозы).
Ацетил-КоА поступает из митохондрий в цитоплазму при помощи цитратного механизма. В митохондриях ацетил-КоА взаимодействует с оксалоацетатом (фермент – цитратсинтаза), образующийся цитрат переносится через митохондриальную мембрану при помощи специальной транспортной системы. В цитоплазме цитрат реагирует с HS-КоА и АТФ, вновь распадаясь на ацетил-КоА и оксалоацетат, который затем восстанавливается до малата и уходит оратно вматрикс (фермент – цитратлиаза).
Начальной реакцией синтеза жирных кислот является карбоксилирование ацетил-КоА с образованием малонил-КоА.
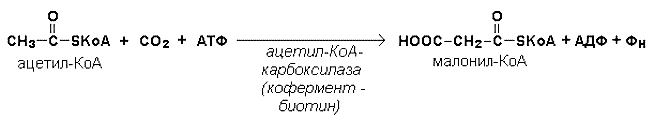
Затем ацетил-КоА и малонил-КоА взаимодействуют с SH-группами ацилпереносящего белка

Далее происходит их конденсация, декарбоксилирование и восстановление образовавшегося продукта

Рис. Реакции одного цикла биосинтеза жирных кислот.
Продукт реакции взаимодействует с новой молекулой малонил-КоА и цикл многократно повторяется вплоть до образования остатка пальмитиновой кислоты.
Биоэнергетика синтеза: n=3,5m-6=3,5*16-6=50 молей АТФ
n- количество молей АТФ, затраченных на синтез пальмитиновой кислоты
m- число углеродных атомов жирной кислоты
|




