Мы поможем в написании ваших работ!
ЗНАЕТЕ ЛИ ВЫ?
|
Характеристика высокоэнергетических фосфатов. Цикл атф-адф
Содержание книги
- Биологические функции белков. Взаимодействие с лигандами.
- Конформация белковых молекул (вторичная и третичная структуры). Активный центр белков. Связывание лигандов.
- Основы кинетики ферментативного катализа
- Кофакторы ферментов. Коферментные функции витаминов группы В.
- Ингибиторы активности ферментов
- Классификация и номенклатура ферментов
- Наследственные (первичные) энзимопатии: нарушение обмена при алкаптонурии, фенилкетонурии, гипераммониемии.
- Активаторы и ингибиторы ферментов
- Регуляция ферментов путем их фосфорилирования — дефосфорилирования
- Процесс включает следующие основные этапы.
- III. Исключение праймеров. Завершение формирования отстающей цепи ДНК
- Биосинтез рнк (транскрипция). Посттранскрипционные модификации рнк
- Биосинтез белков. Основные компоненты и этапы этого процесса. Посттрансляционный процессинг белков.
- Регуляция экспрессии генов. Теория оперона. Индукция и репрессия синтеза белков.
- Молекулярные механизмы генетической изменчивости. Мутации. Наследственные протеинопатии.
- Молекулярные механизмы генных, хромосомных и геномных мутаций
- Биологический код, его свойства, значение в биосинтезе белка. Взаимодействие кодонов с антикодонами
- Макроэргические соединения. Их роль в клетке
- Характеристика высокоэнергетических фосфатов. Цикл атф-адф
- Тема 5. 6. Разобщение дыхания и синтеза атф
- Глк-глюкоза, Фру-фруктоза, Гал-галактоза
- Синтез глюкозы (глюконеогенез)
- Биосинтез и мобилизация гликогена: последовательность реакций, регуляция гормонами, физиологическое значение.
- Регуляция активности фосфорилазы гормонами.
- Основные пути превращения глюкозы в печени.
- Регуляция обмена углеводов инсулином, глюкагоном, адреналином, кортизолом.
- Пентозофосфатный путь окисления глюкозы, физиологическое значение.
- Гликолипиды и гликопротеины. Представления о строении и функциях их углеводных компонентов.
- I. Структура, классификация и свойства основных липидов организма человека
- Б. Структура и классификация фосфолипидов и сфинголипидов
- Пищевые жиры: норма суточного потребления, переваривание, всасывание продуктов.
- В поддержании гомеостаза холестерола в организме. Биохимия желчнокаменной болезни
- Тема 8. 3. Хиломикроны - транспортная форма экзогенных жиров
- Бета-Окисление жирных кислот. Последовательность реакций. Энергетическое значение.
- Механизмы биосинтеза жирных кислот. Регуляция этого процесса.
- Регуляция синтеза жирных кислот.
- Биосинтез жиров в печени и жировой ткани. Регуляция синтеза жиров
- Депонирование и мобилизация жиров в жировой ткани. Механизм регуляции активности липазы гормонами.
- Фосфолипиды, строение, биологическая роль.
- Холестерин, этапы биосинтеза, биологические функции, регуляция биосинтеза.
- Транспортные формы холестерина. Нарушения транспорта. Семейная гиперхолестеринемия. Атеросклероз.
- Причины и факторы риска развития атеросклероза
- Первичные эндогенные формы истощения
- Представления о биосинтезе фосфолипидов. Липотропные вещества.
- Связь между обменом белков и углеводов
- Влияние инсулина, глюкагона и адреналина на обмен жиров.
- Тема 9. 2. Переваривание белков в желудке и кишечнике, всасывание аминокислот
- Реакции трансаминирования, ферменты, их коферментная группа. Биологическое значение реакций. Определение аминотрансфераз с диагностической целью
- Окислительное дезаминирование аминокислот. Глутаматдегидрогеназа. Значение этой реакции.
- Непрямое дезаминирование аминокислот: последовательность реакций, ферменты, биологическое значение.
В живых организмах существует целая группа органических фосфатов, гидролиз которых приводит к освобождению большого количества свободной энергии. Такие соединения называют высокоэнергетическими фосфатами (табл. 6-2).
Как видно из табл. 6-2, разные фосфорили-рованные соединения обладают разным запасом свободной энергии. К группе высокоэнергетических фосфатов, помимо АТФ, относят енолфосфаты, ангидриды и фосфогуанидины. Соединения, расположенные в нижней части таблицы, составляют группу низкоэнергетических фосфатов. Центральное место среди этих соединений занимает АТФ (рис. 6-2).
Таблица 6-2. Свободная энергия гидролиза некоторых органических фосфатов
 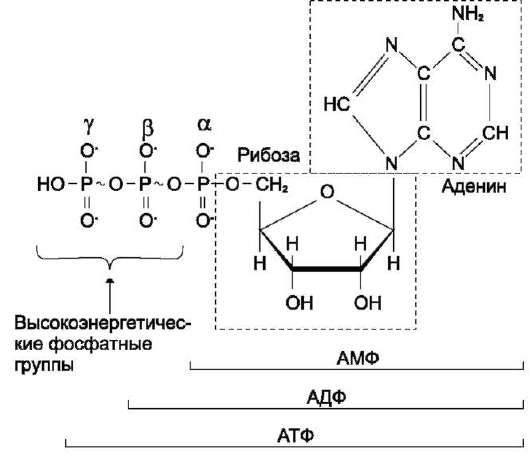
Рис. 6-2. Аденозинтрифосфорная кислота (АТФ).
В молекуле АТФ две высокоэнергетические (макро-эргические) связи β и γ; они обозначены на рисунке знаком ~ (тильда).
АТФ - молекула, богатая энергией, поскольку она содержит две фосфоангидридные связи (β, γ). При гидролизе концевой фосфоангидридной связи АТФ превращается в АДФ и ортофосфат Pi. При этом изменение свободной энергии составляет -7,3 ккал/моль. При условиях, существующих в клетке в норме (рН 7,0, температура 37?C), фактическое значение ΔG0' для процесса гидролиза составляет около -12 ккал/моль. Величина свободной энергии гидролиза АТФ делает
возможным его образование из АДФ за счёт переноса фосфатного остатка от таких высокоэнергетических фосфатов, как, например, фосфоенолпируват или 1,3-бисфосфоглицерат; в свою очередь, АТФ может участвовать в таких эндергонических реакциях, как фосфорилирова-ние глюкозы или глицерина. АТФ выступает в роли донора энергии в эндергонических реакциях многих анаболических процессов. Некоторые биосинтетические реакции в организме могут протекать при участии других нуклеозидтрифос-фатов, аналогов АТФ; к ним относят гуанозин-трифосфат (ГТФ), уридинтрифосфат (УТФ) и цитидинтрифосфат (ЦТФ). Все эти нуклеотиды, в свою очередь, образуются при использовании свободной энергии концевой фосфатной группы АТФ. Наконец, за счёт свободной энергии АТФ совершаются различные виды работы, лежащие в основе жизнедеятельности организма, например, такие как мышечное сокращение или активный транспорт веществ.
Таким образом, АТФ - главный, непосредственно используемый донор свободной энергии в биологических системах. В клетке молекула АТФ расходуется в течение одной минуты после её образования. У человека количество АТФ, равное массе тела, образуется и разрушается каждые 24 ч.
Использование АТФ как источника энергии возможно только при условии непрерывного синтеза АТФ из АДФ за счёт энергии окисления органических соединений (рис. 6-3). Цикл АТФ-АДФ - основной механизм обмена энер-
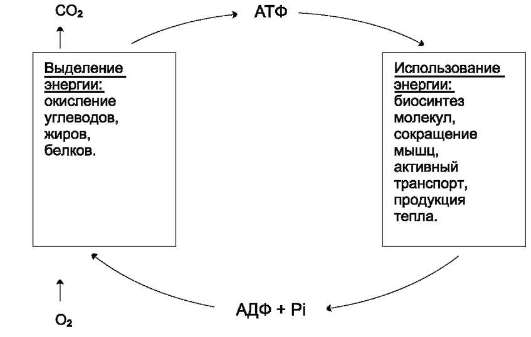
Рис. 6-3. Цикл АТФ-АДФ.
гии в биологических системах, а АТФ - универсальная «энергетическая валюта».
29) НАД-зависимые дегидрогеназы, механизм действия. Важнейшие субстраты НАД-зависимых дегидрогеназ.
НАД-зависимые дегидрогеназы. В качестве кофермента содержат НАД и НАДФ. Пиридиновое кольцо никотинамида способно присоединять электроны и протоны водорода.
НАД-зависимая дегидрогеназа расположена на матриксной поверхности внутренней мембраны митохондрий отдает пару электронов водорода на ФМН-зависимую дегидрогеназу. При этом из матрикса пара протонов переходит также на ФМН и в результате образуется ФМН Н2. В это время пара протонов, принадлежащих НАД выталкивается в межмембранное пространство.
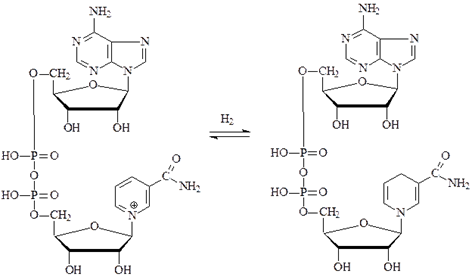
Субстратами НАД-зависимых дегидрогеназ являются спирты, альдегиды, амины, дикарбоновые и кетокислоты. Все НАД-зависимые дегидрогеназы являются анаэробными дегидрогеназами, то есть посылают атомы водорода на ближайший в окислительной цепи другой фермент.
Примером НАД-зависимой дегидрогеназы является алкогольдегидрогеназа из печени животных. Фермент состоит из двух субъединиц, каждая из которых несет молекулу НАД+ и атом Zn. Он катализирует реакцию окисления спирта в альдегид:
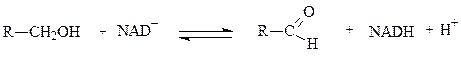
|