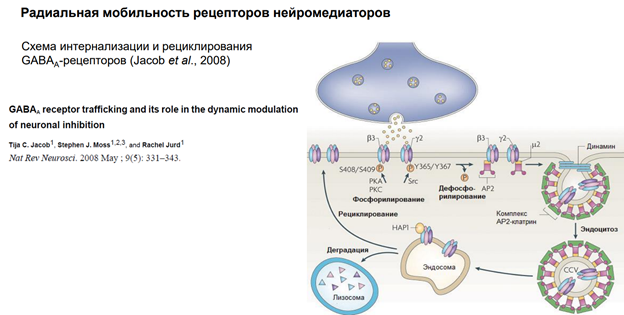
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Посттетаническая потенциацияСодержание книги
Поиск на нашем сайте
Эти процессы давно известны. Это статья статья 1965 года. Эксперимент был выполнен на цилиарном ганглии цыплёнка. Есть входы в цилиарный ганглий, которые мы стимулируем одиночным импульсом и получаем ВПСП (волна вверх на рисунке справа). А – в покое – ВПСП маленький. Добавили тут Д-тубокурарин, чтобы он немного снизил активность, поскольку он блокирует проводимость (иначе были бы очень большие эффекты) Затем мы потенциируем мембрану, т.е прикладываем высоко частотную стимуляцию (50 раздражений в секунду в течение 20 сек). Т.е. на вход цилиарного ганглия мы подаем 50*20=1000 стимулов. И после этого спустя 15 сек даем единичный стимул. Фрагменты Б-Ф – после тетануса. Получаем увеличенный потенциал. *Электротонический потенциал – потенциал в пресинаптическом волокне; он электротонически проникает в постсинаптический нейрон; и его амплитуда одна и та же, не меняется со временем (это означает, что активация входа одна и та же, активируем в одной и той же степени его). Но мембрана постсинаптическая потенциирована, видим ВПСП, котор уменьшается со временем. Если повысить концентрацию Кальция и дать 500 стимулов, то длительность потенциации увеличится до 2 ч.
Если отразить предыдущие картинки (фигура справа) на графике, получим вот такую кривулину. По Оу - амплитуда в %, по Ох – время в мин. За 100% (или за единицу) принимается нормативный потенциал до воздействия. Потом потенциируем. В норме – первая точка слева. Тетанус обозначен вертикальной заштрихованной областью на оси Ох. После тетануса практически в 8 раз идет увеличение амплитуды. Это очень существенное увеличение, оно явно не функционально. Фронт быстро нарастает и затихает к 10-ой минуте. Но не возвращается…на картинке, по крайней мере, выходит на плато. Скорее всег отвернётся через большее время в норму.
Есть процесс, за счет которого может происходить десенситизация, когда нейрон возвращается в исходное свое положение. Десенситизация была хорошо описана на ацетилхолиновых рецепторах. *Десенситизацию можно и на нервно-мышечной передаче можно увидеть. *чувствительность=афинность не изменится (на слайде не верно написано), т.к чувствительность – это степень связывания. Что происходит? На примере ацетилхолиновых рецепторов. В закрытом состоянии нейромедиатора нет. Под действием ацх открытое состояние, наблюдаем все катионные токи: вход Натрия и Кальция. А если будем дальше продолжать стимуляцию, канал перестанет пропускать катионы («адаптируется»). Если сравнить с потенциал зависимыми Натриевыми каналами, то можно считать, что эта десенситизация соответствует инактивации каналов. Но нейромедиатор остается в канале (см рис самого правого канала). Фактически инактивированнй канал открыт, связан с ацх, но тока нет(!), т.е можно сказать, что он полуоткрыт. За счет чего это происходит? Один из механизмов инактивации – фосфорилирование субъединиц со стороны цитоплазмы. Это некая защитная реакция рецептора.
Тут мышца была погружена в Кальциевый раствор. Нужно просто уменьшить вероятность экзоцитоза на стимуляцию, и, естественно, более низкая концентрация Кальция. Это нервно-мышечный препарат. Смотрим рисунок А. Тут изображена потенциация синапса. Каждый последующий стимул в пачке высокочастотный повышает амплитуду. Если прекратим потенциацию и через некоторе вермя дадим одиночный стимул, получим одиночный пик слегка увеличенной амплитудой. Рисунок Б. Если окунем в курара. Курара – блокатор. Дадим высокую концентрацию Кальция, получим депрессию. Понятное дело, по мере срабатывания каналов, они все больше закрываются курарином, получаем искусственную депрессию. Рисунок С. Если суммировать два этих эффекта, получим, что в первые моменты на ритмическую стимуляцию получим потенциацию. Через некоторое время дадим одиночный стимул, получим депрессию.
Причины депрессии: 1) истощение синапт пузырьков, т.к если кальция и так мало, а мы его потратим на синаптич передачу. 2) (см слайд), например, модуляция выброса ацетилхолина … Т.е мы можем подействовать на пресинаптич теримнали через Р2 рецепторы. И через них можно получить обратную регуляцию снижения экзоцитоза (ретроградная регуляция выброса медиатора другим медиатором). АТФ – известна нам как источник энергии (одна из главных функций в нервной системе), но в то же время у него есть нейромдиаторная функция через соответсвующие рецепторы. И АТФ содержится практически во всех везикулах, куда закачан нейромедиатор (закон/ принцип Деила). Принцип Деила: в везикулах может быть более чем 1 молекула нейромедиатора/ полипептиды и т.д
|
||
|
|
Последнее изменение этой страницы: 2022-01-22; просмотров: 127; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.103 (0.01 с.) |