Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Свойства сенсорных рецепторовСодержание книги
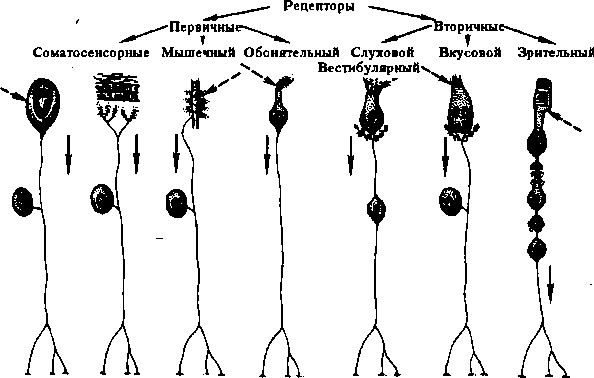
Поиск на нашем сайте Возбудимость рецепторов очень высока, она превышает чувствительность новейших технических аппаратов, регистрирующих соответствующие сигналы. В частности, для возбуждения фоторецептора сетчатки глаза достаточно 1—2 квантов света, для обонятельного рецептора — одной молекулы пахучего вещества. Однако возбудимость вис- церорецепторов ниже, чем экстероре- цепторов. Низкую возбудимость имеют болевые рецепторы, приспособленные к ответу на действие повреждающих раздражителей. Адаптация рецепторов— это уменьшение их возбудимости при длительном действии раздражителя, выражающееся в снижении амплитуды РП и, как следствие, частоты импульсации в афферентном нервном волокне. На начальном этапе действия раздражителей важную роль в адаптации рецепторов могут играть их вспомогательные структуры. Например, быстрая адаптация рецепторов вибрации (тельца Пачини) обусловлена тем, что их капсула пропускает к нервному окончанию только быстро изменяющиеся параметры раздражителя и «отфильтровывает» его статические составляющие. Следует заметить, что термин «темновая адаптация» фоторецепторов означает повышение их возбудимости. Одним из механизмов адаптации рецепторов является накопление Са2+ в нем при возбуждении, что активирует Са2+-зависимые калиевые каналы; выход К+ через эти каналы из клетки препятствует деполяризации ее мембраны и, следовательно, формированию РП. Обнаружены биохимические реакции, блокирующие формирование РП. Значение адаптации рецепторов заключается в том, что она защищает организм от избыточного потока импульсов, иногда — от неприятных ощущений. Спонтанная активность некоторых рецепторов (фоно-, вестибу- ло-, термо-, хемо- и проприорецепторы) без действия на них раздражителя, что связано с проницаемостью клеточной мембраны для ионов, приводящей периодически к снижению ПП до КП и генерации ПД в нервном волокне. Возбудимость таких рецепторов выше, чем рецепторов без фоновой активности, — даже слабый раздражитель способен значительно повысить частоту импуль- сации нейрона. Фоновая активность рецепторов в условиях физиологического покоя участвует в поддержании тонуса ЦНС и бодрствующего состояния организма. Функцией сенсорных рецепторов (лат. sensus — чувство, receptum — принимать) является восприятие раздражителей — изменение внешней и внутренней среды организма. Это осуществляется с помощью преобразования энергии раздражения в РП, который обеспечивает возникновение нервных импульсов. Каждый вид рецепторов в процессе эволюции приспособлен к восприятию одного или нескольких видов раздражителей. Такие раздражители называются адекватными. К ним рецепторы имеют наибольшую чувствительность (например, рецепторы сетчатки глаза возбуждаются при действии 1—2 квантов световой энергии). К другим — неадекватным раздражителям — рецепторы малочувствительны. Неадекватные раздражители также могут возбудить сенсорные рецепторы, однако энергия этих раздражителей должна быть в миллионы и миллиарды раз больше энергии адекватных. Сенсорные рецепторы являются первым звеном в рефлекторном пути и периферической частью сенсорных систем. Классификация сенсорных рецепторов проводится по нескольким критериям (рис. 4.1).
По скорости адаптации различают быстро адаптирующиеся (фазные), медленно адаптирующиеся (тонические) и смешанные (фазнотонические) рецепторы, адаптирующиеся со средней скоростью. Примером быстро адаптирующихся рецепторов являются рецепторы вибрации (тельца Пачини) и прикосновения (тельца Мейснера) кожи. К медленно адаптирующимся рецепторам относятся проприорецепторы, часть болевых рецепторов, механорецепторы легких. Со средней скоростью адаптируются фоторецепторы сетчатки глаза, терморецепторы кожи. В зависимости от вида воспринимаемого раздражителя выделяют четыре типа рецепторов, а именно: хеморецепторы — вкусовые и обонятельные рецепторы, часть сосудистых и тканевых рецепторов (реагирующих на изменение химического состава крови, лимфы, межклеточной жидкости) — имеются в гипоталамусе (например, в пищевом центре) и продолговатом мозге (дыхательный центр); механорецепторы — расположены в коже и слизистых оболочках, опорнодвигательном аппарате, сосудах, внутренних органах, слуховой, вестибулярной и тактильной сенсорных системах; терморецепторы (их подразделяют на тепловые и холодовые) — находятся в коже, сосудах, внутренних органах, различных отделах ЦНС (гипоталамусе, среднем, продолговатом и спинном мозге); фоторецепторы — расположены в сетчатке глаза, воспринимают световую (электромагнитную) энергию. В зависимости от способности воспринимать один или более видов раздражителей выделяют моносенсорные (обладают максимальной чувствительностью к одному виду раздражителей, например рецепторы сетчатки) и полисенсорные (воспринимают несколько адекватных раздражителей, например механический и температурный или механический, химический и болевой) рецепторы. Примером являются ирритантные рецепторы легких, болевые рецепторы. По расположению в организме рецепторы делят на экстеро- и интерорецепторы. К интерорецепторам относятся рецепторы внутренних органов (висцерорецепторы), сосудов и ЦНС. Разновидностью интерорецепто- ров являются рецепторы опорно-двигательного аппарата (проприорецепторы) и вестибулярные рецепторы. К экстеро- рецепторам относятся рецепторы кожи, видимых слизистых оболочек (например, слизистой рта) и органов чувств: зрительные, слуховые, вкусовые, терморецепторы, обонятельные. По ощущениям рецепторы подразделяют на зрительные, слуховые, вкусовые, обонятельные терморецепторы, тактильные, болевые (ноцицепторы) — это свободные нервные окончания, которые имеются в зубах, коже, мышцах, сосудах, внутренних органах. Они возбуждаются при действии механических, термических и химических (гистамин, брадикинин, К+, Н+ и др.) раздражителей. Механизм возбуждения рецепторов. При действии адекватного раздражителя в первичном рецепторе возникает РП, представляющий собой деполяризацию клеточной мембраны обычно вследствие движения ионов Na+ внутрь клетки. РП — это локальный потенциал, он является раздражителем нервного окончания (за счет своего электрического поля) и обеспечивает возникновение
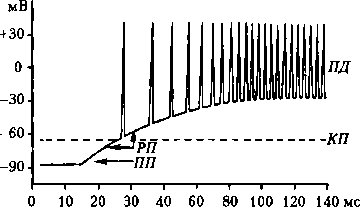
Рис. 4.2. Типичные взаимоотношения между амплитудой РП и частотой ПД, возникающих в афферентном нервном волокне при сверхпороговых уровнях РП (по А. Гайтон, 1985)
ПД в мякотных волокнах — в первом перехвате Ранвье, в безмякотных — в непосредственной близости от рецептора. Во вторичных рецепторах при действии раздражителя сначала также возникаем РП в рецепторной клетке вследствие движения Na+ в клетку (вкусовые и обонятельные рецепторы) или К+ (слуховые и вестибулярные рецепторы). Под влиянием РП в синаптическую щель выделяется медиатор, который, действуя на постсинаптическую мембрану, обеспечивает формирование ГП (тоже локальный). Последний является раздражителем (электрическое поле), обеспечивающим возникновение ПД в нервном окончании, как и в окончаниях с первичными рецепторами. Зависимость частоты ПД в афферентном нервном волокне от величины РП и ГП показана на рис. 4.2. 4.2. Физиология нервных волокон Структурно-функциональная характеристика нервных волокон. Нервные волокна — это отростки нейронов, с помощью которых осуществляется связь между нейронами и с исполнительными клетками. Имеется два типа нервных волокон: миелинизированные и немиелинизированные (безмиелино- вые). Оболочку безмиелино- вых волокон образуют шванновские клетки (леммоциты), в которые погружаются осевые цилиндры нервных волокон. Оболочку миелинизированных волокон образуют в периферической нервной системе также шванновские клетки (миелоциты, глиальная клетка), формирующие миелин (многослойная обертка мембран миелоцитов — до 100), а в ЦНС — олигодендроциты (глиальные клетки, т.е. те же шванновские клетки). Миелиновая оболочка через равные участки (около 1 мм) прерывается, образуя свободные от миелина небольшие участки — перехваты Ранвье. Основную часть миелина (78 % сухого веса) составляют липиды, обеспечивающие изолирующие свойства оболочки. Нервные волокна обеспечивают п р о в е д е н и е возбуждения и аксонный транспорт, выполняющий трофическую функцию нейрона. По классификации Дж. Эрлангера и X. Гассера выделяют три типа волокон: А, В и С (см. таблицу). Волокна типов А и В являются миелинизированными. А-волокна — это афферентные и эфферентные волокна соматической нервной системы; к волокнам типа В принадлежат преганглионарные волокна ВНС. С- волокна немиелинизированные — это постганглионарные волокна ВНС, а также афферентные волокна от некоторых болевых, тепловых и висцеральных рецепторов. Аксонный транспорт. Основная масса веществ образуется в трофическом центре нейрона, расположенном преимущественно около ядра, а использу- Таблица
|
|||
|
|
Последнее изменение этой страницы: 2021-04-12; просмотров: 219; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.150 (0.011 с.) |