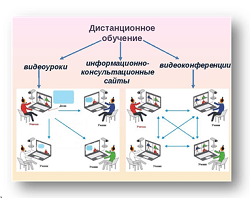
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Клеточная теория.клетка-элементарная еденица живого.клетки прокариот и эукариот.увеличение числа клеток.гомологичность в строении клеток.многоклеточный организм-сложный ансамбль клеток.Содержание книги
Поиск на нашем сайте ЭПС, характеристика,ультраструктура,функции. Рибосомы, строение и роль в синтезе белка. ЭПС встречается практически у всех эукариот. Выделяют 2 типа ЭР: гранулярный(шероховатый) и гладкий. Гранулярный ЭП: Представлен замкнутыми мембранами,которые образуют на сечениях вытянутые мешки,цистерны или же имеют вид узких каналов. Со стороны гиалоплазмы эти мембраны покрыты мелкими темными,почти круглыми частицами,гранулами. Эти гранулы являются рибосомами,связанными с мембранами ЭР.На мембранах рибосомы расположены в виде полисом,имеющим вид плоских спиралей,розеток или гроздей. Гранулярный ЭР является важным местом синтеза белков. Количество рибосом на ЭР четко связано с его синтетической активностью.Рибосомы в составе полисом гиалоплазмы синтезируют белки,необходимые для собственно клеточных нужд,для поддержания и обеспечения данной клетки.Рибосомы же связанные с мембранами ЭР, учавствуют в синтезе белка,выводимых из данной клетки.А также эти рибосомы учавствуют в процессе сегрегации,обособления этих белков,в их изоляции от основных функционирующих белков клетки. Было обнаружено,что гранулярный ЭР может синтезировать интегральные и периферические белки мембран и даже часть белков матрикса цитоплазмы. Гладкий (агранулярный) ЭР Часть мембраной ретикулярной системы. Он также представлен мембранными,образующими мелкие вакуоли и трубки,канальца,которые могут ветвица,сливаться друг с другом. На мембранах гладкого ЭР нет рибосом. Было установлено непрерывность перехода между гладкой формой ЭР и гранулярной его формой или гранулярной в гладкую. Гладкий ЭР является вторичным по отношению к гранулярному ЭР. Деятельность гладкого ЭР можно связать с метаболизмом липидов и некоторых внутриклеточных полисахаридов. Гладкий ЭР особенно в большом количестве встречается в клетках секретирующих стероиды, в клетках печени,в мышечных волокнах. Среди высших растений гладкий ЭР встречается в клетках тканей,учавствующих в синтезе и транспорте терпенов,стероидов и липидов. Рибосомы присутствуют и в прокариотических и в эукариотических клетках. В клетках эукариот есть 2 разновидности рибосом:рибосомы собственно цитоплазмы и рибосомы локализованные в таких клеточных органоидах,как митохондриях и хлоропластах. Рибосомы прокариот имеют коффициент седементации 70S(S- константа седементации). У рибосом цитоплазмы эукариот коффициент седементации 80S.Рибосомы митохондрий 70S,но они сложнее. Митохондриальные хромосомы разных групп эукариот отличаются по коффициенту седементации.Структурная организация рибосом всех групп принципиально одинакова.Рибосома состоит из 2 суедениц: большой и малой.У рибосом 70S прокариот эти субеденицы имеют коффициент седементации соответственно 50 и 30 S,у рибосом 80S-60 и 40S.В клетках существует динамическое равновессие между субеденицами (70 S взаимно переходит 50+30S)Нетранслирующие рибосомы постояно обмениваются субеденицами. Непосредственная сборка рибосом идет лишь в момент работы. Динамическое равновесие между суеденицами можно изменить вправо или влево,изменяя содержание ионов магния(происходит их диссоциация).Субеденицы рибосом состоят из РНК и белка.РНК слогает карказ, к которому крепятся белки. Функция рибосом:белковый синтез. В процессе инициации и-РНК присоеденяется к малой субеденице, потом к ним ассоциирует большая субеденица. Синтез АТФ. Структура АТФ-синтетазного комплекса, локализация, принцип функционирования. Гипотеза П. Митчелла. Строении АТФсинтетазы. Мембраны несущие в своем составе элементы переноса электронов и сопряженног с ним синтеза АТФ – называются сопряженными и представлены 2 слоями белков и 1 липидов. Синтез АТФ осуществляется АТФ-синтетазным комп. Дых. Цепт – АТФ-с – дых.цепь. АТФ-с. -,Дых. цепь АТФ синтетаза - состоит из многочисленных субединиц и из 2 ух комплексов 1 – полностью погружен в мембрану (ножка гриба), а, в, с, d. 2 - сверху (шляпка) 5 типов субед. Глобулярные ά β Вытянутые ε γ δ. Суб. Ед. разные функции: β - активные центры в них синтез АТФ, ά - субстрат связывающие, γ δ.- контакт, ε - ингибитор, с - протонный канал. С протонныц канал через который идет поток н+ на субединицу бетта. Бета является центром синтеза АТФ,на нее поступает АДФ плюс фосфат (Фн), на них образуется макроэргическая связь – АТФ. Хемиосматическая гипотеза П. Митчелла -1961 г. Та энергия, которая необходима создается за счет перекачки электронов из матрикса митох. в цитоплазму, в результате создается трансмембранный градиент ∆μН+, - универсальная форма запаса энергии в клетке. Транс мембранный градиент состоит из разности потенциалов и концентрации протонов. При функционировании Д\Ц все время происходит перекачка протонов и образуется и разность потенциало и разность концентрации протонов. ∆μН+, -энергия кот. Создает Д\цю – мембрана заряжена. Две формы запаса энергии АТФ и ∆μН+, но они различны и нужны обе. ∆μН+, - связана в основном с мембранами преимущественно, Ферменты как биокатализаторы, их спецефичность и механизм действия. Изоферменты. Регуляция активности обычных и аллостерических ферментов. Ретроингибирование. Влияние физико-химических факторов на активность ферментов. Относит. молекул. масса белков, облад-х ферментными св-ми, колеблется от 15 тыс. до н-х млн. Ферменты явл-ся глобулярными белками, вкл-т простые (однокомпонентные) и сложные (двукомпонентные). Относит. молекул. масса белков, облад-х ферментными св-ми, колеблется от 15 тыс. до н-х млн. Ферменты явл-ся глобулярными белками, вкл-т простые (однокомпонентные) и сложные (двукомпонентные).
(E-фермент, К-кофермент, AH2,В- субстраты).Соединение белковой части с небелковой осущ-тся за счет ионных, водородных связей, гидрофобных взаимодействий, реже- с помощью ковалентных связей. Функции: 1) участие в акте катализа; 2) осуществление контакта между ферментативным белком и суб-стратом; 3) стабилизация апофермента. Апофермент определяет специфичность действия. Один и тот же апофермент может функционировать в составе разных ферментов. НАД является коферментом многих дегидрогеназ. Ферменты внутри клетки содержатся и действуют в определенных ее орга В ядрах ферменты синтеза нуклеиновых кислот ДНК-полимеразы, РНК- полимеразы). В лизосомах гидролитические ферменты. В пероксисомах ферменты метаболизма гликолевой кислоты и утилизации пероксида водорода. В матриксе митохондрий-ферменты окисления жирных кислот. В строме хлоропластов-рибулозодифосфат карбоксилаза и др.ферменты, участвующие в синтезе углеводов из СО2. В мембраны хлоропластов встроены АТФ-азы и переносчики электронов, функционирующие в процессе фотосинтеза. При ферментативном катализе наблюдается снижение активации. Реакции протекают ступенчато. Активационный барьер разбивается на несколько более низких. 3 стадии: 1)присоединение молекулы субстрата S к ферментату Е; 2)превращение субстрата; 3)определение конечных прод-в р-ции от фермента. 1-ая стад. -быстрая-образование ферментных комплексов, 2-ая-медленная. Образование ферментсубстратного комплекса возможно из-за определенного сродства ферментов к субстрату. Различают положительные эффекторы (метаболиты, вызывающие аллостерическую активацию (обратимое связывание ферментами некоторых метаболитов вызывает уменьшение или увеличение активности фермента). И различают отрицательные, связывание которых с ферментом снижает скорость реакции благодаря аллостерическому ингибированию. Аллостерический эффектор связывается с аллостерическим центром, изменяет конформацию фермента, что вызывает изменение сродства фермента к субстрату. Если в системе накопления много конечного продукта, то его образование будет ингибировано, в результате чего конечный продукт как ингибитор первого фермента в метаболическом цикле, ведущем к синтезу этого продукта. Это ретронгибирование рН, при котором скорость реакции максимальна, называется оптимумом, при отклонении рН в любую сторону от этого значения скорость реакции снижается. При нагревании выше 80 С преобладающее большинство ферментов полностью денатурирует. Температурный оптимум 40-60 С. Большинство ферментов хорошо сохраняет активность при положительных и отрицательных температурах.Только холодолабильные инактивируются при понижении t от 30 до 40 градусов.
Клеточная теория.клетка-элементарная еденица живого.клетки прокариот и эукариот.увеличение числа клеток.гомологичность в строении клеток.многоклеточный организм-сложный ансамбль клеток. Клеточная теория-это обобщенные представления о строении клеток как едениц живого,об их размножении и роли в формировании многоклеточных организмов. заслуга швана: 1-клетка-элементарная еденица живого 2-клетки разных организмов гомологичны по своему строению 3-размножение клеток происходит путем деления исходной клетки 4-многоклеточные организмы представляют собой сложный ансамбль клеток, объединенные в целостные интегрированные системы тканей и органов, подчиненных и связанных между собой межклеточными, гуморальными и нервными формами регуляции. Живому свойствен ряд совокупных признаков, таких как способность к воспроизведению (репродукции),использовании и трансформации энергии, метаболизм, чувствительность, изменчивость. Такую совокупность этих признаков можно обнаружить на клеточном уровне. Клетка как таковая обладает всеми вместе взятыми свойствами, отвечающими определению живое. Клетки делят на прокариотические и эукариотические. Содержимое прокариотной клетки одето плазматической мембраной, играющей роль активного барьера между собственно цитоплазмой клетки и внешней средой.обычно снаружи есть еще и клеточная стенка или оболочка-продукт клеточной активности. У прокариотической клетки нет морфологически выраженного ядра, но присутствует в виде так называемого нуклеоида-зона заполненная ДНК. В матриксе цитоплазмы прокариот клетки располагаются многочисленные рибосомы, цитоплазматические мембраны же обычно выражены не так сильно,как у эукариотических клеток. Эукариотические клетки, как правило, больше прокариотических. В них есть ядро. В цитоплазме существует целый набор специфических структур, органелл, выполняющих отдельные специфические функции: мембранные структуры-система ЭПС, аппарат Гольджи, лизосомы, митохондрии, пластиды, центриоли. Наличие мембранных структур-микротрубочки,микрофиламенты и тд. Общие черты прокариотической и эукариотической клеток: обе одеты плазматической мембраной, синтез белка у них происходит на рибосомах, сходны синтез РНК и репликация ДНК, похожи и биоэнергетические процессы. Клетка-это ограниченная активность мембраной, упорядоченная, структурированная система биополимеров (белков, НК) и их макромолекулярных комплексов, участвующих в единой совокупности метаболических и энергетических процессов, осуществляющих поддержание и воспроизведение всей системы в целом. Гомологичность клеток-сходство по коренным свойствам и отличие по второстепенным. Все клетки организмов растительного и животного происхождения сходны, гомологичны. Гомология определяется тем, что клеточные функции можно грубо подразделитьна 2 группы: обязательные и факультативные. Обязательные функции направлены на поддержание жизнеспособности самих клеток, осуществляются специальными внутриклеточными структурами. Прокариотические клеткимогут отличаться друг от друга толщиной и устройством клеточной стенки, складчатостью плазматической мембраны и тд. Но общий план строения прокариотической клетки остается постоянный. Это же наблюдается и у эукариотической клетки. Схожи в строении животные и растительные клетки. У всех эукариотических клеток всегда есть ядро. Строение и функции внутриклеточных структур также похожи по строению у всех эукариотических клеток. Такое сходство определяется гомологичностью общеклеточных функций, связанных с поддержанием самой системы. Но есть большое разнообразие клеток в пределах одного многоклеточного организма (нервные, мышечные).Различия клеток связана со специализацией их функций, с развитием особых клеточных аппаратов. Так в мышечной клетке покроме основных структур, есть в большом количестве фибриллярные элементы. Гомологичность в строении клеток определяется сходством общеклеточных функий, направленных на поддержание жизни самих клеток и на их размножение. Разнообразие же в строении клеток-это результат функциональной специализации или в следствии эволюционной приспособленности, изменчивости. Размножение клетоке прокариотич. И эукариот. Происходит только путем деления исходной клетки, которому предшествует воспроизведение ее генетического материала (редупликация ДНК). Прокар. клетка делится бинарным образом, простой перегородкой, без участия каких-либо специальных аппаратов деления. У эукар.кл. способом деления является митоз (или мейоз). При этом образуется специальный аппарат деления-клеточное веретено. Другая форма клеточного деления-амитоз, или простое деление-может встречаться в ряде паталогических случаев или при делении прлиплоидных ядер. Организм человека развивается всего из 1 клетки, зиготы, содержит более 100 различных типов клеток. Индивидуальное развитие от 1 кл. до многоклеточного зрелого организма-результат последовательного избирательного включения работы разных генных участков хромосом в различных клетках. Это приводит к дифференцировке клеток.
|
||
|
|
Последнее изменение этой страницы: 2017-02-19; просмотров: 337; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.217.151 (0.006 с.) |