Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Полимеразная цепная реакция (Дейвис, 1990)Содержание книги
Поиск на нашем сайте Для проведения ПЦР готовили необходимое количество реакционной смеси для анализа необходимого числа проб в пропорциях, представленных в таблице 1. Для устранения погрешности, вносимой дозаторами при смешивании микрообъемов, общую смесь готовили из расчета на одну пробу больше, чем необходимо. Таблица 1 Состав реакционной смеси для проведения ПЦР
Смесь тщательно перемешивали на миксере «Rotamix RM-1» (ELMI, Латвия), кратко центрифугировали в микроцентрифуге «Eppendorf Centrifuge 5414» при 14 тыс. об/мин. В маркированные пробирки объемом 0.5 мл вносили по 23 мкл полученной реакционной смеси и по 2 мкл ДНК, полученной в результате реакции обратной транскрипции. Компоненты перемешивали на миксере «Rotamix RM-1» (ELMI, Латвия), смесь кратко центрифугировали в микроцентрифуге «Eppendorf Centrifuge 5414» при 14 тыс. об/мин. Для предотвращения испарения жидкости реакционную смесь покрывали двумя каплями минерального масла. Реакционную смесь инкубировали в амплификаторе «Терцик» («ДНК-Технология», Россия) по программе Fas Hot Start. Условия инкубации реакционной смеси в амплификаторе «Терцик»:
94ºС – 30с 65ºС – 30с 45 циклов 72ºС – 40с для однонуклеотидного полиморфизма Fas -1377 (A/G):
67,5ºС – 40с 45 циклов 72ºС – 30с Для выявления полиморфизмов были использованы праймеры, нуклеотидная последовательность которых показана в таблицах 2 и 3. Также указана первичная структура праймеров для амплификации промоторного участка гена Fas (таблица 4).
Таблица 2 Структура праймеров для амплификации промоторного участка гена Fas, содержащего олигонуклеотидный полиморфизм Fas-670A\G
Таблица 3 Структура праймеров для амплификации промоторного участка гена Fas, содержащего олигонуклеотидный полиморфизм Fas-1377A\G
Таблица 4 Первичная структура праймеров для амплификации промоторного участка гена Fas, содержащего олигонуклеотидный полиморфизм Fas-670A\G
Результаты реакции анализировали методом электрофореза в агарозном геле.
2.4 Визуализация результатов ПЦР методом электрофореза нуклеиновых кислот в 1,5% агарозном геле (Maniatis, 1982) Готовили 1,5%-ный агарозный гель с лунками для внесения проб. Для этого 1,5 г агарозы разводили в 100 мл рабочего трис-ацетатного буферного раствора (24,2 г трис-(гидрокси)аминометана, 5,7 мл ледяной уксусной кислоты, 10 мл 0,5 М раствора этилендиаминотетрауксусной кислоты, pH 8.0) и добавляли 10мкл 10%-ного водного раствора бромида этидия. Суспензию нагревали на водяной бане до полного растворения агарозы, периодически помешивая. Раствору давали остыть примерно до 50˚С и заливали в форму для агарозного блока, вставляли гребенку для образования лунок и давали агарозе застыть до состояния геля. Из-под слоя масла отбирали 10 мкл раствора амплифицированной ДНК и смешивали с 10 мкл буферного раствора для нанесения проб (50% глицерина, 0,25% бромфенолового синего в дистиллированной воде). Полученную смесь переносили в лунки геля. Камеру для электрофореза «Helicon» (г. Москва) подключали к источнику питания «Эльф-2» (Россия), соблюдая полярность. Проводили электрофорез в течение 15 минут при напряжении 200 вольт (краситель должен пройти не менее 2 см геля). Отключали ток. Помещали гель в окно трансиллюминатора «Biokom UVT-1» (Франция). Результат регистрировали с использованием системы видеодокументирования «VITRan». Статистическую обработку полученных результатов проводили с помощью пакета программ Statistica v. 8.0. Данные представляли в виде относительных и абсолютных частот. Анализ различий частот признаков в группах проводили с использованием критерия % и двустороннего теста точного критерия Фишера. Различия считали статистически значимыми при р<0.05.
Результаты и их обсуждение По данным литературы известны ассоциации частот генотипов ОНП Fas-670A/G ОНП Fas-1377A/G в промоторной части гена Fas с различными заболеваниями. Среди населения, проживающего на территории Российской Федерации, не проводилось исследований по выявлению частот генотипов ОНП Fas-670A/G ОНП Fas-1377A/G в промоторной области гена Fas, что определяет практическую значимость данного исследования. Анализ аллельных вариаций в составе ОНП Fas -670A/G, Fas -1377G/A проводился методом аллель-специфической ПЦР с помощью олигонуклеотидных праймеров. Продукты реакции детектировали с помощью электрофореза в 1.5 % агарозном геле, содержащем флуоресцентный краситель бромид этидия. На первом этапе настоящего исследования проводилось генотипирование образцов периферической крови здоровых доноров, больных раком толстого кишечника и герпесвирусной инфекцией по ОНП Fas-670A/G. Результаты исследований генотипов ОНП Fas-670A/G представлены в таблице 5.
Таблица 5 Частота генотипов однонуклеотидного полиморфизма Fas-670A/G в клетках периферической крови больных раком толстого кишечника, герпесвирусной инфекцией и здоровых доноров
* - статистически значимые различия по сравнению со здоровыми донорами (р<0,05) У здоровых доноров частота генотипа FAS-670AA составляла 3,9%, генотипа FAS-670AG –60,8%, генотипа FAS-670GG –35,3%. Генотип FAS-670AG встречался у всех 44 больных раком толстого кишечника (100%). У больных герпесвирусной инфекцией частота генотипа FAS-670AA составляла 8,6%, генотипа FAS-670AG – 77,1%, генотипа FAS-670GG – 14,3%. Исследовали частоты генотипов Fas-670A/G в клетках периферической крови больных раком толстого кишечника, герпесвирусной инфекцией и здоровых доноров. Было показано, что у больных раком толстого кишечника частота генотипа -670AG статистически значимо (в 1,65 раза) была выше по сравнению со здоровыми донорами (р<0,001). При этом у больных герпесвирусной инфекцией частота генотипа -670GG была в 2,5 раза ниже (p=0,034) (рис.3).
Рис.3. Частота генотипов Fas-670A/G в клетках периферической крови больных раком толстого кишечника и герпесвирусной инфекцией. * - статистически значимые различия (р<0,05) по сравнению со здоровыми донорами Исследовали также частоты генотипов ОНП Fas-670A/G у мужчин и женщин, больных раком толстого кишечника, герпесвирусной инфекцией и у здоровых доноров. Результаты исследований представлены в таблице 6.
Таблица 6 Частота генотипов однонуклеотидного полиморфизма Fas-670A/G в клетках периферической крови мужчин и женщин, больных раком толстого кишечника, герпесвирусной инфекцией и здоровых доноров
* - статистически значимые различия по сравнению со здоровыми донорами (р<0,05)
При сравнении фрагмента генома мужчин, больных раком толстого кишечника и герпесвирусной инфекцией, со здоровыми не выявлены статистически значимые различия по частотам как гомозигот AА и GG, так и гетерозигот AG по ОНП Fas-670A/G (рис.4).
Рис.4. Частота генотипов Fas-670A/G в клетках периферической крови мужчин, больных раком толстого кишечника и герпесвирусной инфекцией. По сравнению со здоровыми донорами у женщин, больных раком толстого кишечника, частота генотипа -670AG статистически значимо (в 1,83 раза) была выше (р<0.001). При этом генотипы -670АА и -670GG в образцах крови онкологических больных не выявлялись (рис.5).
Рис.5. Частота генотипов Fas-670A/G в клетках периферической крови женщин, больных раком толстого кишечника и герпесвирусной инфекцией. * - статистически значимые различия (р<0,05) по сравнению со здоровыми женщинами При сравнении мужчин и женщин, больных раком толстого кишечника, а также мужчин и женщин, больных герпесвирусной инфекцией, и здоровых мужчин и женщин не было обнаружено статистически значимых различий по частотам как гомозигот AА и GG, так и гетерозигот AG по ОНП Fas-670A/G. На следующем этапе настоящего исследования проводилось генотипирование образцов периферической крови здоровых доноров, больных раком толстого кишечника и герпесвирусной инфекцией ОНП Fas-1377AG. Результаты представлены в таблице 7.
Таблица 7 Частота генотипов однонуклеотидного полиморфизма Fas-1377A/G в клетках периферической крови больных раком толстого кишечника, герпесвирусной инфекцией и здоровых доноров
* - статистически значимые различия по сравнению со здоровыми донорами (р<0,05) У здоровых доноров частота генотипа FAS-1377AA составляла 2,2%, генотипа FAS-1377AG –67,4%, генотипа FAS-1377GG –30,4%. Генотип FAS-1377AA встречался у 22,2% больных раком толстого кишечника, генотип FAS-1377AG – у 41,7%, генотип FAS-1377GG – у 36,1%. У больных герпесвирусной инфекцией частота генотипа FAS-1377AA составляла 13,8%, генотипа FAS-1377AG – 20,7%, генотипа FAS-1377GG – 65,5%. Было выявлено, что при сравнении со здоровыми донорами у больных раком толстого кишечника частота генотипа Fas-1377AG статистически значимо была ниже в 1,62 раза (р=0,03). В отношении других генотипов статистически значимых различий обнаружено не было; по сравнению со здоровыми донорами у больных герпесвирусной инфекцией частота генотипа Fas-1377GG статистически значимо (в 2,2 раза) была выше (p=0,004). Наряду с этим генотип -1377АG детектировался реже в 3,26 раза, чем у здоровых доноров (р=0,002) (рис.6).
Рис.6. Частота генотипов Fas-1377A/G в клетках периферической крови больных раком толстого кишечника и герпесвирусной инфекцией. * - статистически значимые различия (р<0,05) по сравнению со здоровыми донорами Исследовали также частоты генотипов ОНП Fas-1377A/G у мужчин и женщин, больных раком толстого кишечника и герпесвирусной инфекцией, и у здоровых доноров. Результаты исследований представлены в таблице 8.
Таблица 8 Частота генотипов однонуклеотидного полиморфизма Fas-1377A/G в клетках периферической крови мужчин и женщин, больных раком толстого кишечника, герпесвирусной инфекцией, и здоровых доноров
* - статистически значимые различия по сравнению со здоровыми донорами (р<0,05) По сравнению со здоровыми донорами у мужчин, больных герпесвирусной инфекцией, частота генотипа -1377АG детектировалась на реже в 2,96 раза (р = 0,016) (рис.7).
Рис.7. Частота генотипов Fas-1377A/G в клетках периферической крови мужчин, больных раком толстого кишечника и герпесвирусной инфекцией. * - статистически значимые различия (р<0,05) по сравнению со здоровыми мужчинами По сравнению со здоровыми донорами у женщин, больных раком толстого кишечника, частота генотипа Fas-1377AG статистически значимо (в 1,67 раза) была ниже (р=0,03). В отношении других генотипов статистически значимых различий обнаружено не было. При сравнении со здоровыми донорами у женщин, больных герпесвирусной инфекцией, частота генотипа Fas-1377GG статистически значимо (в 2,5 раза) была выше (p=0,01). Наряду с этим генотип -1377АG детектировался в 6,5 раз реже, чем у здоровых доноров (р=0,004) (рис.8).
Рис.8. Частота генотипов Fas-1377A/G в клетках периферической крови женщин, больных раком толстого кишечника и герпесвирусной инфекцией. * - статистически значимые различия (р<0,05) по сравнению со здоровыми женщинами При сравнении мужчин и женщин, больных раком толстого кишечника, герпесвирусной инфекцией и здоровых мужчин и женщин не было обнаружено статистически значимых различий по частотам как гомозигот AА и GG, так и гетерозигот AG по ОНП Fas-1377A/G. Полученные результаты показали, что некоторые частоты генотипов однонуклеотидных полиморфизмов -670A/G и -1377G/A у больных раком толстого кишечника и герпесвирусной инфекцией отличаются от соответствующих значений частот генотипов здоровых доноров, что может свидетельствовать о влиянии наличия соответствующих аллельных вариантов генотипов на предрасположенность к данным заболеваниям. Кроме этого, были выявлены статистически значимые различия частот некоторых исследованных генотипов у мужчин и женщин с данными патологиями, что позволяет предположить о том, что наличие исследованных генотипов по-разному влияет на предрасположенность к данным заболеваниям у мужчин и женщин. У пациентов с герпесвирусной инфекцией частоты генотипов Fas -670A/G не отличались от частот соответствующих генотипов здоровых доноров. В то же время у всех больных раком толстого кишечника был выявлен только генотип FAS-670AG, что в 1,65 раза выше по сравнению со здоровыми донорами. Частоты генотипов FAS-670AG, выявленные в нашем исследовании, отличались от частот соответствующих генотипов, представленных в литературе (Hofflman et al., 2009), где частоты генотипов были более равномерными между собой (FAS -670AA 24,2%, FAS -670AG – 46,3%, FAS -670GG – 29,5%), что, с одной стороны, отражает особенности частот генотипов в разных популяциях, а, с другой стороны, может являться особенностью данной патологии. Не было выявлено различий между частотами генотипов FAS-670AG у мужчин и женщин с раком толстого кишечника. Тем не менее, было показано, что по сравнению со здоровыми донорами, у женщин, больных раком толстого кишечника, частота генотипа FAS-670AG выше в 1,83 раза при этом у мужчин таких особенностей выявлено не было. Данные говорят о различном влиянии наличия генотипов FAS-670AG у мужчин и женщин на предрасположенность к данной патологии. У больных раком толстого кишечника частоты генотипов FAS-1377A/G не отличались от здоровых доноров. У больных герпесвирусной инфекцией частота генотипа Fas-1377 GG была в 2,2 раза выше по сравнению со здоровыми донорами, при этом у мужчин частота генотипа -1377АG была в 2,96 раза ниже по сравнению со здоровыми донорами, а у женщин, больных герпесвирусной инфекцией, частота генотипа Fas-1377GG в 2,5 раза выше, чем у здоровых доноров. Таким образом, в отличие от рака толстого кишечника, частота генотипов FAS-1377A/G у больных герпесвирусной инфекцией различна как по сравнению со здоровыми донорами, так и у мужчин и женщин по сравнению друг с другом. Выводы 1. Выявлено, что у здоровых доноров частота генотипа FAS-670AA составляла 3,9%, генотипа FAS-670AG –60,8%, генотипа FAS-670GG –35,3%; частота генотипа FAS-1377AA составляла 2,2%, генотипа FAS-1377AG –67,4%, генотипа FAS-1377GG –30,4%. 2. Установлено, что генотип FAS-670AG встречался у всех 44 больных раком толстого кишечника (100%). Генотип FAS-1377AA встречался у 22,2% больных раком толстого кишечника, генотип FAS-1377AG – у 41,7%, генотип FAS-1377GG – у 36,1%. По сравнению со здоровыми донорами у больных раком толстого кишечника частота генотипа -670AG статистически значимо в 1,65 раза была выше. 3. Показано, что у больных герпесвирусной инфекцией частота генотипа FAS-670AA составляла 8,6%, генотипа FAS-670AG – 77,1%, генотипа FAS-670GG – 14,3%.; частота генотипа FAS-1377AA составляла 13,8%, генотипа FAS-1377AG – 20,7%, генотипа FAS-1377GG – 65,5%. При сравнении со здоровыми донорами у больных герпесвирусной инфекцией частота генотипа Fas-1377GG статистически значимо в 2,2 раза была выше. 4. При сравнении фрагмента генома мужчин, больных раком толстого кишечника и герпесвирусной инфекцией, со здоровыми не выявлены статистически значимые различия по частотам как гомозигот AА и GG, так и гетерозигот AG по ОНП Fas-670A/G. У мужчин, больных герпесвирусной инфекцией, частота генотипа -1377АG детектировалась реже в 2,96 раза в сравнении со здоровыми донорами. 5. По сравнению со здоровыми донорами у женщин, больных раком толстого кишечника, частота генотипа -670AG статистически значимо в 1,83 раза была выше. У женщин, больных герпесвирусной инфекцией, частота генотипа Fas-1377GG статистически значимо в 2,5 раза выше, чем у здоровых доноров. Цитированная литература 1. Барышников А.Ю., Шишкин Ю.В. Иммунологические проблемы апоптоза. – М.: Эдиториал УССР, 2002. – 320с. 2. Льюин Б. Гены: Пер. с англ.- М.:Мир,-1987-544с 3. Фогель Ф., Мотульски А. Генетика человека: в 3-х т. Т.2. Пер. с англ.- М.: Мир-1990-378с 4. Arasteh J.M., Sarvestani E.K., Aflaki E., Amirghofran Z. Fas gene polymorphisms in systemic lupus erythematosus and serum levels of some apoptosis-related molecules. // Int. J. Gynecol. Cancer. 2006. Vol.16. P. 179-182. 5. Arnold P. What are Single Nucleotide Polymorphisms? // Int. J. Cancer. 2008. Vol. 103. P. 221-225. 6. Bel Hadj Jrad B. A polymorphism in FAS gene promoter associated with increased risk of nasopharyngeal carcinoma and correlated with anti-nuclear autoantibodies induction. // Cancer Lett. 2006. Vol. 233. P. 21-27. 7. Ben Aleya W., Sfar I., Mouelhi L. et al. Association of Fas/Apo1 gene promoter (-670 A/G) polymorphism in Tunisian patients with IBD. // World J. Gastroenterol. 2009. Vol. 15, №29. P. 3643-3648. 8. Cao Y., Miao X.P., Huang M.Y. et al. Polymorphisms of death pathway genes FAS and FASL and risk of nasopharyngeal carcinoma. // Mol. Carcinog. 2010. Vol. 49, №11. P. 944-950. 9. Crew K.D. et al. Genetic polymorphisms in the apoptosis-associated genes FAS and FASL and breast cancer risk. // Carcinogenesis. 2007. Vol. 28. P. 2548-2551. 10. Dybikowska A., Sliwinski W., Emerich J., Podhajska A.J. Evaluation of Fas gene promoter polymorphism in cervical cancer patients. // Int J. Mol. Med. 2004. Vol. 14, №3. P. 475-478. 11. Erdogan M. et al. Fas/Fas ligand gene polymorphism in patients with papillary thyroid cancer in the Turkish population. // J. Endocrinol. Invest. 2007. Vol. 30. P. 411-416. 12. Farre L., Bittencourt A.L., Silva-Santos G. et al. Fas 670 promoter polymorphism is associated to susceptibility, clinical presentation, and survival in adult T cell leukemia. // Virus Res. 2012. Vol. 163, №1. P. 178-182. 13. Gormus U. et al. Fas-1377 A/G polymorphism in lung cancer. // In Vivo. 2007. Vol. 21. P. 663-666. 14. Gormus U. et al. Fas-1377A/G and FasL-844 T/C gene polymorphisms and epithelial ovarian cancer. // Anticancer Res. 2007. Vol. 27. P. 991-994. 15. Hsu P.I. et al. Polymorphisms of death pathway genes FAS and FASL and risk of premalignant gastric lesions. // Anticancer Res. 2008. Vol. 28. P. 97-103. 16. Huang Q.R., Morris D., Manolios N. Identification and characterization of polymorphisms in the promoter region of the human Apo-1/Fas (CD95) gene. // Mol. Immunol. 1997. Vol. 34. P. 577–582. 17. Ikehara S.K. et al. A polymorphism of C-to-T substitution at -31 IL1B is associated with the risk of advanced gastric adenocarcinoma in a Japanese population. // J. Hum. Genet. 2006. Vol. 51. P. 927-933. 18. Kanemitsu S., Ihara K., Saifddin A. et al. A functional polymorphism in Fas (CD95/APO-1) gene promoter associated with systemic lupus erythematosus. // J. Rheumatol. 2002. Vol. 29. P. 1183–1188. 19. Koshkina N.V. et al. Exploratory analysis of Fas gene polymorphisms in pediatric osteosarcoma patients. // J. Pediatr. Hematol. Oncol. 2007. Vol. 29. P. 815-821. 20. Krippl P., Langsenlehner U., Renner W. et al. Polymorphisms of Death Pathway Genes FAS and FASL in Esophageal Squamous-Cell Carcinoma. // J. Nat. Cancer Inst. 2004. Vol. 96, №19. Р. 1478-1479. 21. Lai H.C. et al. Genetic polymorphisms of FAS and FASL (CD95/CD95L) genes in cervical carcinogenesis: an analysis of haplotype and gene-gene interaction. // Gynecol. Oncol. 2005. Vol. 99. P. 113-118. 22. Lai H.C. et al. Single nucleotide polymorphism at Fas promoter is associated with cervical carcinogenesis. // Int. J. Cancer. 2003. Vol. 103. P. 221-225. 23. Lei D., Sturgis E. M., Wang L.-E. et al. FAS and FASLG Genetic Variants and Risk for Second Primary Malignancy in Patients with Squamous Cell Carcinoma of the Head and Neck. // Cancer Epidemiol. Biomarkers Prev. 2010. Vol. 19. P. 1484-1491. 24. Li C. et al. Functional polymorphisms in the promoter regions of the FAS and FAS ligand genes and risk of bladder cancer in south China: a case-control analysis. // Pharmacogenet. Genomics. 2006. Vol. 16. P. 245-251. 25. Li C. et al. Polymorphisms of the FAS and FAS ligand genes associated with risk of cutaneous malignant melanoma. // Pharmacogenet. Genomics. 2006. Vol. 16 P. 253-263. 26. Lima L. et al. Association between FAS polymorphism and prostate cancer development. // Prostate Cancer Prostatic Dis. 2008. Vol. 11. P. 94-98. 27. Lima L., Calais da Silva F. M., Calais da Silva F. E. et al. Fas-670 polymorphism and prostate cancer. // J. Clin. Onc. 2007. Vol.25, №18. Р. 21028. 28. Marín L.A., Muro M., Moya-Quiles M.R. et al. Study of Fas (CD95) and FasL (CD178) polymorphisms in liver transplant recipients. // Tissue Antigens. 2006. Vol. 67, №2. P. 117-126. 29. Melville B. What are ОНП? // Eur. J. Hum. Genet. 2000. Vol. 8, №2. Р. 154-165. 30. Nelson H.H. et al. Fas/APO-1 promoter polymorphism is not associated with non-melanoma skin cancer. // Cancer Epidemiol. Biomarkers Prev. 2001. Vol. 10. P. 809-810. 31. Norrgard K., Schultz J. et al. Natural selection has driven population differentiation in modern humans. // Nature Genetics. 2008. Vol. 40. P. 340-345. 32. Park S.H. et al. Polymorphisms in the FAS and FASL genes and risk of lung cancer in a Korean population. // Lung Cancer. 2006. Vol. 54. P. 303-308. 33. Pengfei S., Qi D., Chao Q.et al. Functional polymorphisms in cell death pathway genes FAS and FAS ligand and risk of prostate cancer in a Chinese population. // The Prostate. 2011.Vol. 71, №10. P. 1122–1130. 34. Sibley K. et al. Functional FAS promoter polymorphisms are associated with increased risk of acute myeloid leukemia. // Cancer Res. 2003. Vol. 63. P. 4327-4330. 35. Smith K., Lancia G., Lippert R., Schwartz R. ОНП Problems, Complexity and Algorithms. // LNCS. 2002. Vol. 2161. P. 182-193. 36. Sun T. et al. FASL -844C polymorphism is associated with increased activation-induced T cell death and risk of cervical cancer. // J. Exp. Med. 2005. Vol. 202. P. 967-974. 37. Sun T. et al. Polymorphisms of death pathway genes FAS and FASL in esophageal squamous-cell carcinoma. // J. Natl. Cancer Inst. 2004. Vol. 96. P. 1030-1036. 38. Sziller I., Nguyen D., Halmos A. et al. An A>G polymorphism at position −670 in the Fas (TNFRSF6) gene in pregnant women with pre-eclampsia and intrauterine growth restriction. // Mol. Hum. Reprod. 2005. Vol.11, №3. Р. 207-210. 39. Tamandani D.M. et al. Association of Fas-670 gene polymorphism with risk of cervical cancer in North Indian population. // Clin. Exp. Obstet. Gynecol. 2008. Vol. 35. P. 183-186. 40. Ter-Minassian M. et al. Apoptosis gene polymorphisms, age, smoking and the risk of non-small cell lung cancer. // Carcinogenesis. 2008. Vol. 29. P. 2147-2152. 41. Ueda M. et al. Fas gene promoter -670 polymorphism (A/G) is associated with cervical carcinogenesis. // Gynecol. Oncol. 2005. Vol. 98. P. 129-133. 42. Ueda M. et al. Fas gene promoter -670 polymorphism in gynecological cancer. // Int. J. Gynecol. Cancer. 2006. Vol. 16. P. 179-182. 43. Vallinoto A.C., Santana B.B., Dos Santos E.L. et al. FAS -670A/G single nucleotide polymorphism may be associated with human T lymphotropic virus-1 infection and clinical evolution to TSP/HAM. // Eur. J. Gynaecol. Oncol. 2009. Vol. 30, №2. P. 142-144. 44. Wang L.E. et al. Fas A670G polymorphism, apoptotic capacity in lymphocyte cultures, and risk of lung cancer. // Lung Cancer. 2003. Vol. 42. P. 1-8. 45. Yang M. et al. Functional variants in cell death pathway genes and risk of pancreatic cancer. // Clin. Cancer Res. 2008. Vol. 14. P. 3230-3236. 46. Zhang B., Sun T., Xue L. et al. Functional polymorphisms in FAS and FASL contribute to increased apoptosis of tumor infiltration lymphocytes and risk of breast cancer. // Carcinogenesis. 2007. Vol. 28, №5. P. 1067-1073. 47. Zhang H. et al. Importance of FAS-1377, FAS-670, and FASL-844 polymorphisms in tumor onset, progression, and pigment phenotypes of Swedish patients with melanoma: a case-control analysis. // Cancer J. 2007. Vol. 13. P. 233-237. 48. Zhang X., Miao X., Sun T. et al. Functional polymorphisms in cell death pathway genes FAS and FASL contribute to risk of lung cancer. // J. of Med. Genet. 2005. Vol. 42, №6. P. 479-484. 49. Zhang Z. et al. Polymorphisms of FAS and FAS ligand genes involved in the death pathway and risk and progression of squamous cell carcinoma of the head and neck. // Clin. Cancer Res. 2006. Vol. 12. P. 5596-5602. 50. Zhang Z., Xue H., Gong W. et al. FAS promoter polymorphisms and cancer risk: a meta-analysis based on 34 case-control studies.// Carcin. 2009. Vol. 30, №3. Р.487-493. 51. Zhou R.M., Wang N., Chen Z.F. et al. Polymorphisms in promoter region of FAS and FASL gene and risk of cardia gastric adenocarcinoma. // J. Gastroenterol. Hepatol. 2010. Vol. 25, №3. P. 555-561. 52. Zhu Q., Wang T., Ren J. et al. FAS-670A/G polymorphism: A biomarker for the metastasis of nasopharyngeal carcinoma in a Chinese population. Clin. Chim. Acta. 2010. Vol. 411, №3, 4. P.179-183. 53. Zoodsma M. et al. Interleukin-10 and Fas polymorphisms and susceptibility for (pre)neoplastic cervical disease. // Int. J. Gynecol. Cancer 2005. Vol. 15. P. 282-290. 54. Zucchi F., da Silva I.D., Ribalta J.C. et al. Fas/CD95 promoter polymorphism gene and its relationship with cervical carcinoma. // Immunol. Invest. 2010. Vol. 39, №1. P. 27-38.
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2017-01-26; просмотров: 280; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.153 (0.009 с.) |

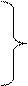 для однонуклеотидного полиморфизма Fas -670 (A/G):
для однонуклеотидного полиморфизма Fas -670 (A/G):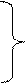 94ºС – 30с
94ºС – 30с