Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Внутрішньоклітинне окиснення продуктів гідролітичного розщеплення жирівСодержание книги
Поиск на нашем сайте Гліцерин і вищі жирні кислоти, що утворилися в процесі травлення жирів, а також у результаті розщеплення три-гліцеридів у жирових депо за участі тканинних ліпаз, зазнають подальших перетворень. У клітинах різних органів і тканин вони окиснюються до кінцевих продуктів (С02 і Н20) або в процесі окиснення використовуються для біосинтезу інших сполук. Окиснення гліцерину. Процес окиснення гліцерину починається з фосфорилювання його під дією ферменту гліцеролкінази. Донатором фосфатної кислоти в цій реакції виступає АТФ. У результаті утворюється гліцеринфосфатна кислота:
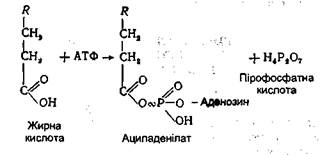
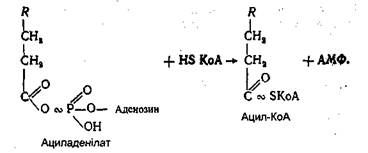
Найбільш інтенсивно фосфорилювання гліцерину відбувається в клітинах печінки. Гліцеринфосфатна кислота, що утворилася, за допомогою гліцеринфосфатдегідрогенази перетворюється в діоксіацетонфосфат, який ізомерізується в гліцеральдегід-3-фосфат. Останній окислюється до фосфогліцеринової кислоти подібно до того, як це відбувається в процесі гліколізу. Поряд з цим проміжні продукти окиснення гліцерину можуть використовуватися організмом для біосинтезу жирів, фосфогліцеридів і вуглеводів. Оквснення вищих жирних кислот. Всі жирні кислоти перед окисненням підлягають активації. Цей процес також відбувається головним чином у печінці і здійснюється за участю АТФ і коензиму А. Схематично цей процес можна подати таким чином: спочатку жирна кислота взаємодіє з АТФ з утворенням ациладенілату.
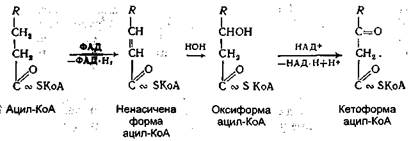
Ацил-КоА далі вступає на шлях окиснення, який інтенсивно відбувається в мітохондріях. Весь процес окиснення ацил-КоА полягає в постійному дегідруванні його з відщепленням від ацил-КоА жирної кислоти двовуглецевих фрагментів у вигляді ацетил-КоА (активної форми оцтової кислоти). Оскільки весь процес дегідрування супроводжується відщепленням від жирної кислоти двовуглецевих фрагментів у β-положенні, він має назву α-окиснення. Дегідрування ацил-КоА відбувається під дією специфічних дегідрогеназ за участю коферментів ФАД і НАД+:
Кетоформа ацилкоензиму А за допомогою ще однієї молекули HS-KoA ферментативним шляхом розщеплюється на дві молекули - ацетил-КоА й ацил-КоА:
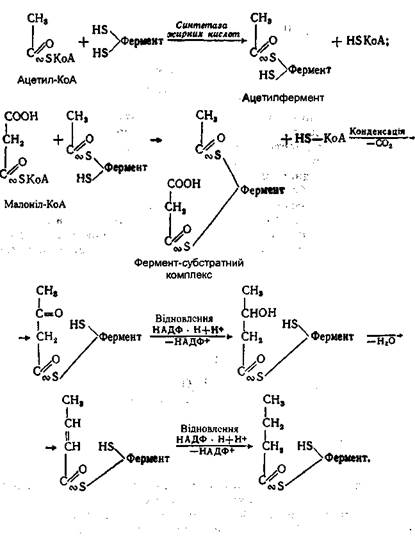
Скоротившись на два вуглецеві атоми, молекула ацил-КоА знову піддається дворазовому дегідруванню з відщепленням нової молекули ацетил-КоА. Зрештою вся молекула вищої жирної кислоти розщеплюється до молекул ацетил-КоА. Якщо жирна кислота мала парну кількість атомів вуглецю, то наприкінці утворюється ціле число молекул ацетил-КоА. Так, молекула стеаринової кислоти (C17H35COOH) утворює 9 молекул, пальмітинова (С15Н31СООН) - 8 молекул ацетил-КоА. Утворені молекули аце-тил-КоА "згорають" у циклі Кребса уже відомим нам шляхом. При розщепленні вищих жирних кислот до молекул ацетил-КоА звільняється близько 30% енергії, інші 70% виділяються при окисненні ацетил-КоА до СO2 і Н2O в циклі Кребса. Розглянемо як приклад енергетичний баланс окиснення однієї молекули пальмітинової кислоти, яка утворює 8 молекул ацетил-КоА. Така кількість ацетил-КоА утворюється в результаті семи послідовних відщеплень, кожне з яких двічі супроводжується дегідруванням: один раз при участі ФАД, другий - за участі НАД. За рахунок кожної утвореної відновленої форми ФАД • Н2 і НАД Н + Н+ у дихальному ланцюзі утворюється 5 молекул АТФ {ФАД Н2 - 2 молекули і НАД • Н + Н+ - 3). Усього ж це становить 35 молекул, тону що відщеплення ацетил-КоА відбулося 7 разів (це майже стільки ж, скільки утворюється при окисненні молекули глюкози до СО2 і Н2О). За рахунок окиснення кожної молекули ацетил-КоА в циклі Кребса утворюється ще 12 молекул АТФ, а в результаті окиснення 8 молекул - 96 молекул АТФ. Таким чином, окиснення однієї молекули пальмітинової кислоти супроводжується утворенням 131 молекули АТФ (35 + 96). Враховуючи одну молекулу АТФ, використану для активації пальмітинової кислоти, "чистий прибуток" АТФ складе 130 молекул, в яких акумульовано 4353-5442 кДж енергії. Оскільки в складі тригліпериду міститься три залишки вищої жирної кислоти, а окиснення гліцерину також супроводжується утворенням АТФ, загальний підсумок окиснення однієї молекули тригліцериду буде ще більшим. БІОСИНТЕЗ ТРИГЛІЦЕРИДІВ Синтез нейтральних жирів складається з трьох процесів: утворення вищої жирної кислоти, утворення гліцерину І сполучення цих речовин у молекулу триглідериду. Синтез вищих жирних кислот. Місцем утворення вищих жирних кислот є цитоплазма. У процесі беруть участь активна форма оцтової кислоти, тобто ацетил-КоА, і вуглекислий газ. На першому етапі біосинтезу при взаємодії ацетил-КоА і СО2 утворюється проміжна сполука - малоніл-КоА. Утворення цієї сполуки відбувається за участі вітаміну Н з використанням енергії АТФ:
У процесі біосинтезу вищої жирної кислоти малоніл-КоА розщеплюється на ацетил-КоА і СО2, а окремі молекули ацетил-КоА з'єднуються між собою в довгий ланцюг. Отже, малоніл-КоА не входить до складу ланцюга вищої жирної кислоти, а служить лише проміжною формою, яка забезпечує утворення вищої жирної кислоти з окремих молекул ацетил-КоА. Весь процес з'єднання молекул ацетил-КоА здійснюється за допомогою ферменту синтетази жирних кислот, яка містить дві сульфгідрильні групи: центральну і периферичну. На центральній тіоловій групі здійснюється реакція конденсації між малоніл-КоА й ацетил-КоА з виділенням СО2 і відновлення утвореного продукту, а периферична група служить для утримання утвореного ланцюга:
У результаті з'єднання двох молекул ацетил-КоА і двох відновних реакцій утворюється фрагмент вищої жирної кислоти, який складається з чотирьох атомів вуглецю. На наступному етапі утворений фрагмент переноситься з центральної тіолової групи ферменту на периферичну:
При цьому центральна тіолова група звільняється і знову вступає в реакцію з новою молекулою малоніл-КоА. Потім у тій же послідовності відбувається реакція конденсації між утвореним ланцюгом і молекулою малоніл-КоА з виділенням С02 І реакції відновлення продукту, що утворився знову. Таким чином, при багаторазовому повторенні цих реакцій вуглеводний ланцюг усе більше подовжується, доки не з'єднає 16-18 атомів вуглецю. Після цього синтезована вища жирна кислота взаємодіє з молекулою коензима А, утворюючи активну форму у вигляді ацил-КоА й звільняючи при цьому синтетазу жирної кислоти:
Біосинтез тригліцеридів. Утворення жиру відбувається в результаті взаємодії молекули гліцерину з трьома молекулами вищих жирних кислот. Основним джерелом гліцерину в організмі є проміжний продукт окиснення вуглеводів - діоксіацетон-монофосфат, який шляхом відновлення перетворюється спочатку в гліцеринфосфатну кислоту, а потім у вільний гліцерин. Субстратом у біосинтезі жирних кислот, як ми тільки-но побачили, служить активна форма оцтової кислоти - ацетил-КоА. Як показали дослідження, гліцерин вступає б реакцію у вигляді гліцеринфосфату, а жирні кислоти - у вигляді своєї активної форми - ацил-КоА. На першому етапі біосинтезу триглі-церидів відбувається утворення фосфатидної кислоти - загального проміжного продукту в біосинтезі жирів І фосфатидів: і Далі фосфатидна кислота ферментативним шляхом розщеплюється на фосфатну кислоту і α, β-дигліцерид, що також є загальним проміжним продуктом у біосинтезі жирів і фосфатидів:
Утворений α, β-дигліцерид взаємодіє з третьою молекулою ацилкоензиму А, утворюючи молекулу тригліцериду:
Синтезований таким шляхом специфічний для організму жир відкладається в жирових депо. Біосинтез лецитину. Лецитин, як відомо, належить до групи фосфатидів. Для його біосинтезу необхідні такі речовини: гліцерин, вищі жирні кислоти, фосфатна кислота й азотиста основа холін. За своєю будовою лецитин деякою мірою нагадує триглі-цериди. Тому багато етапів у біосинтезі лецитину аналогічні етапам біосинтезу нейтральних жирів. Як уже було сказано, загальною проміжною сполукою в біосинтезі фосфатидів і тригліцеридів є α, β-дигліцерид. Саме до цієї проміжної сполуки і приєднуються фосфатна кислота і холін. Відбувається це в такий спосіб. Спочатку холін фосфорилюється за допомогою АТФ з утворенням фосфохоліну:
який далі взаємодіє з цитидинтрифосфатною кислотою (ЦТФ) і перетворюється в цитидиндифосфохолін:
При взаємодії цитидинфосфохоліну з α, β-дигліцерндом утворюється лецитин:
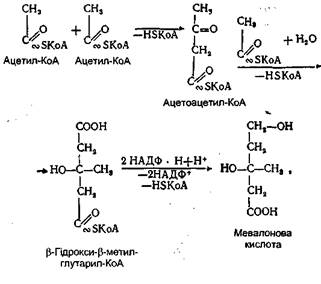
Біосинтез лецитину дуже активно відбувається в стінці тонкої кишки і печінки. Біосинтез холестерину. В організмі людини холестерин синтезується у всіх органах і тканинах. Механізм утворення його дуже складний і довгий час залишався незрозумілим. Завдяки використанню методу мічених атомів удалося встановити всі етапи його синтезу. Вихідною речовиною для синтезу холестерину служить активна форма оцтової кислоти - ацетил-КоА. Окремі етапи його біосинтезу можна схематично передати так: 2 молекули ацетил-КоА конденсуються з утворенням ацетоацетил-КоА. До цієї молекули приєднується третя молекула ацетил-КоА й утворюється складна сполука - (γ-гідрокси-β-метилглутарил-коензим А. З цієї сполуки утворюється мевалонова кислота. Утворення мевалонової кислоти схематично можна показати так:
Мевалонова кислота є одним із найважливіших проміжних продуктів у біосинтезі холестерину. Через ряд проміжних реакцій вона перетворюється в більш складну сполуку. За допомогою АТФ мевалонова кислота перетворюється у свою активну форму - пірофосфат мевалонової кислоти, яка дегідруючись і декарбоксилюючись утворює ізопентілпірофосфат. Після ізомеризації останнього утворюється диметилалілпірофосфат, який здатний уже брати участь у реакціях конденсації і синтезу циклопентанпергідрофенантренового кільця холестерину.
|
||
|
|
Последнее изменение этой страницы: 2016-08-16; просмотров: 860; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.216.150 (0.006 с.) |