Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь FAQ Написать работу КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Концевая шпилька гистоновой мРНКСодержание книги
Поиск на нашем сайте Хотя подавляющее большинство эукариотических мРНК имеют поли(А)-«хвост», мРНК, кодирующие гистоны, синтез которых регулируется клеточным циклом, являются исключением — во всяком случае у млекопитающих и других Metazoa. Вместо поли(А)-«хвоста» на конце таких гистоновых мРНК расположена эволюционно консервативная шпилька с шестью каноническими парами оснований в ее двойной спирали и четырехнуклеотидной петлей на торце (рис. 14.17). Наличие этой структуры, как оказалось, тоже обеспечивает высокий уровень инициации трансляции кэпированной мРНК. В клетках млекопйтающих в составе активных полирибосом был обнаружен небольшой белок, специфически связывающийся с этой шпилькой и необходимый для ее функционирования как усилителя трансляции. В механизме усиления, как и в случае роlу(А) • РАВР, участвуют факторы инициации e1F4G, eIF4E и eIF3, причем для e1F4G показано его прямое взаимодействие с SЕВР. Таким образом, очень вероятно, что SЕВР действует подобно РАВР, формируя белковые мостики SlВР•eIF4G•eIF4E и SLВР•e1F4G•eIF3 между 3'-концом мРНК, с одной стороны, и ее кэпированным 5'-концом и инициирующей 40S рибосомной субъединицей, с другой. Возможно, что РАВР, всегда присутствующий в клетках, тоже присоединяется к eIF4G и участвует в формировании этих мостиков и усилении инициации трансляции.
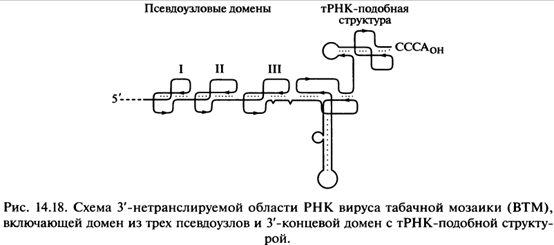
Рис 14.17 Структурированные домены 3’-нетранслируемой области РНК растительных вирусов
Существует группа усилителей трансляции более специального типа — компактные домены, включающие в себя несколько (обычно три) последовательно расположенных псевдоузлов в 3'-НТО, характерная для РНК вирусов растений (в частности, тобамовирусов, тимовирусов и бромовирусов). Эти РНК кэпированы, но не полиаденилированы, и псевдоузловой домен, следующий непосредственно за кодирующей частью РНК, с примыкающей к нему 3'-концевой тРНК-подобной структурой (рис. 14.18) заменяют поли(А)-«хвост». Следует отметить, что РНК некоторых растительных вирусов — например, вируса мозаики люцерны (A1falfa Mosaic Virus, А!МV) — лишены тРНК-подобного домена на конце 3'-НТО и имеют лишь псевдоузловой домен, который выполняет ту же функцию усиления трансляции. Имеются все основания полагать, что именно псевдоузловой домен внутри 3'-НТО вирусных РНК играет основную роль в усилении трансляции предшествующих кодирующих последовательностей. Интересно, что искусственные конструкции мРНК с вирусной 3'-НТО, содержащей псевдоузловые структуры, транслируются в животных клетках или их экстрактах так же эффективно, как мРНК с поли(А)-хвостом. При этом наблюдается функциональный синергизм между вирусной 3'-нетранслируемой областью и кэп-структурой на 5'-конце мРНК. Тот факт, что усиливающий эффект 3'-НТО РНК растительных вирусов проявляется в цитоплазме и экстрактах животных клеток, наводит на мысль о существовании универсального белкового фактора, способного связываться с вышеуказанными структурными элементами «хвостов вирусных РНК и, вероятнее всего, отличного от РАВР. Более детальное изучение функциональной роли различных участков 3'-НТО РНК вируса табачной мозаики показало, что их эффект на трансляцию обусловлен стимуляцией иниииаиии трансляции, и для этого важен прежде всего 72-нуклеотидный домен, расположенный непосредственно после кодирующей последовательности и состоящий из трех псевдоузлов (см. рис. 14.18). Удаление этого псевдоузлового домена приводит к практически полному исчезновению стимулирующей активности всего «хвостам. Избирательное удаление или разрушение одного псевдоузла приводит к снижению стимулирующей активности, но наиболее критичным оказывается удаление третьего, а наименее —первого псевдоузла. У эволюционно и структурно более далеких растительных вирусов в РНК могут присутствовать усилители трансляции совсем другого типа. Так, РНК сател-литного вируса некроза табака и вируса желтой карликовости ячменя не имеют ни поли(А)-хвоста, ни кэп-структуры на 5'-конце, но очень эффективно транслируются благодаря высокой скорости инициации трансляции. Высокая скорость инициации обеспечивается специальными структурированными доменами внутри их 3'-нетранслируемых областей. Было показано, что эти домены функционируют синергично с короткими некэп-пированными лидерами своих РНК; возможно, синергизм достигается за счет взаимодействия ТЕС с лидером — либо прямого (РНК-РНК), либо через посред-ство каких-то белковых факторов.
|
||
|
|
Последнее изменение этой страницы: 2016-08-16; просмотров: 429; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 216.73.217.64 (0.008 с.) |